INTRODUCTION
Stress contributes to symptom exacerbation in many disease states [1], and is closely associated with depression [2,3]. Depression is a major mental health problem and a significant public-health issue in the community. It is caused by chronic physical disease, alcohol addiction, and stress [4,5]. It induces many psychiatric disorders, such as sleep disorder, anxiety, appetite disorder, and in severe cases, suicide. The various brain areas altered by stress include the hippocampus and the dorsal raphe; dorsal raphe nuclei are strongly associated with depression [6]. Depression is closely associated with overactive bladder symptoms [7].
Serotonin, also known as 5-hydroxytryptamine (5-HT), is a monoamine neurotransmitter, which acts as a biochemical messenger and regulator in the brain. 5-HT mediates several important physiological functions including neurotransmission, gastrointestinal motility, hemostasis, and cardiovascular integrity. It is generally thought to be a contributor to feelings of well-being and happiness [8]. 5-HT is closely associated with depression [9], and decrement in 5-HT metabolism was ascertained in the brains of a subgroup of patients with depression [10]. Tryptophan hydroxylase (TPH), the rate-limiting enzyme in 5-HT synthesis, regulates 5-HT activity in the brain [11].
5-HT interacts with 14 serotonin receptors (5-HT receptors) that can be subdivided into 7 classes. Of these, the 5-HT 1A (5-HT1A) receptor is a subtype located in the presynaptic and postsynaptic regions. Presynaptic 5-HT release is regulated by the 5-HT1A autoreceptors in the raphe nuclei of the midbrain. Raphe nuclei are the pacemakers of 5-HT central propagation, which regulates the 5-HT projection in various brain areas [12]. In particular, the dorsal raphe nuclei are closely implicated in mood disorders such as depression [6]. Anxiolytic, antidepressant, and antipsychotic medications act via the activation of 5-HT1A receptors [13,14].
Treatment of stress-induced depression and particularly, its prevention should be geared towards reduction of stress and stress sensitivity [10]. Exercise has been recommended as a valuable strategy for symptom relief in many neuropsychiatric disorders [15-17]. Exercise protects neurons against neuronal injuries and ameliorates the progression of many neuropsychiatric disorders, such as Alzheimer disease, Parkinson disease, and autism [15,16,18,19]. Additionally, exercise is known to alleviate symptoms of depression [20,21]. Exercise also improves sexual function in women taking antidepressants [22].
However, little information is available regarding the effect of treadmill exercise on the stress-induced depressive state. In the present study, we investigated the effect of treadmill exercise on depression induced by stress while focusing on the expression of 5-HT1A receptors in the dorsal raphe.
MATERIALS AND METHODS
Animals
Sprague-Dawley rats, weighing 100±10 g (5 weeks old), were obtained from a commercial breeder (Orient Co., Seoul, Korea) for the experiments. The experimental procedures were performed in accordance with the animal care guidelines of the National Institutes of Health and the Korean Academy of Medical Sciences. The rats were housed under controlled temperature (20°C±2°C) and lighting (7 AM to 7 PM) conditions with food and water available ad libitum. They were randomly divided into 4 groups (n=10 per group): control, control and exercise, stress-induced, and stress-induced and exercise groups.
Induction of Stress
Stress was induced by applying an inescapable 0.2-mA electric foot shock to the rats. One set of electric foot shocks comprised a 6-second shock duration that was repeated 10 times with a 30-second interval. The rats in the stress-induced groups received three sets of electric foot shocks per day. Exposure of the rats to the electric foot shock continued for 7 days.
Exercise Protocol
The rats in the exercise groups were made to run on a motorized treadmill for 30 minutes once a day for 4 weeks. The exercise load consisted of running at a speed of 5 m/min for the first 5 minutes, 8 m/min for the next 5 minutes, and 15 m/min for the last 20 minutes, with a 0° inclination. The rats in the control group and the stress-induced group were left on a treadmill that was not running for the same period as the exercise groups.
Forced Swimming Test
For the confirmation of depressive state, a forced swimming test was performed, according to a previously described method [4,20]. Experimental animals were dropped individually into glass cylinders. The glass cylinders (height, 50 cm; diameter, 15 cm) contained water at a temperature of 27°C and a depth of about 30 cm. All experimental rats underwent a 15-minute pretest for adaptation to the water. One day after the pretest, the animals were tested for 5 minutes. During the test session, immobility time and climbing time were analyzed using a Smart ver. 2.5 video tracking system (Panlab, Barcelona, Spain). Climbing behavior consisted of upward-directed movements of the forepaws along the side of the swim chamber. Immobility was defined to occur when no additional activity was observed other than the actions to keep the animal’s head above the water.
Tissue Preparation
The animals were sacrificed immediately after the forced swimming test. To prepare the brain slices, the animals were fully anesthetized with Zoletil 50 (10 mg/kg, intraperitoneally; Vibac, Carros, France), transcardially perfused with 50mM phosphatebuffered saline (PBS), and then fixed with a freshly prepared solution of 4% paraformaldehyde in 100mM phosphate buffer (PB, pH 7.4). The brains were removed, postfixed in the same fixative overnight, and transferred into a 30% sucrose solution for cryoprotection. Coronal sections of 40-μm thickness were made using a freezing microtome (Leica, Nussloch, Germany).
Immunohistochemistry for 5-HT and TPH Expressions
To visualize 5-HT and TPH expressions, immunohistochemistry for 5-HT and TPH in the dorsal raphe was performed, according to a previously described method [11]. The dorsal raphe spanning 7.20 to 8.00 mm from the Bregma was obtained from each brain. The sections were incubated in PBS for 10 minutes, and then washed three times in the same buffer. The sections were then incubated in 1% H2O2 for 30 minutes. They were then incubated overnight with rabbit anti–5-HT antibody (1:500, Abcam, Cambridge, UK) or rabbit anti-TPH antibody (1:1,000; Oncogene Research Product, Cambridge, UK). This was followed by incubation with biotinylated rabbit secondary antibodies (1:200; Vector Laboratories Inc., Burlingame, CA, USA) for another 1 hour. The secondary antibodies were amplified with the Vector Elite ABC kit (1:100; Vector Laboratories Inc.). Antibody-biotin-avidin-peroxidase complexes were visualized using 0.03% diaminobenzidine (DAB), and the sections were mounted onto gelatin-coated slides. The slides were airdried overnight at room temperature, and coverslips were mounted using Permount (Fisher Scientific, Pittsburgh, PA, USA).
Western Blot for 5-HT1A Receptors
Western blot for the 5-HT1A receptor was performed, according to a previously described method [16,17]. The dorsal raphe was collected and immediately frozen at -70˚C. Protein from each dorsal raphe was extracted. The tissues were homogenized with lysis buffer containing 50mM Tris-HCl (pH 8.0), 150mM NaCl, 10% glycerol, 1% Triton X-100, 1.5mM MgCl2·6H2O, 1mM ethyleneglycotetraacetic acid, 1mM phenylmethylsulfonyl fluoride, 1mM Na2VO4, and 100mM NaF, then ultracentrifuged at 50,000 rpm for 1 hour. Protein content was measured using a Bio-Rad colorimetric protein assay kit (Bio-Rad, Hercules, CA, USA). Protein (30 μg) was separated on sodium dodecyl sulfate-polyacrylamide gels and transferred onto nitrocellulose membranes. Anti-GAPDH antibody (1:3,000; Abcam) and anti–5-HT1A antibody (1:1,000; Abcam) were used as the primary antibodies. Horseradish peroxidase-conjugated antimouse antibodies for GAPDH and 5-HT1A were used as the secondary antibodies. Experiments were performed in normal laboratory conditions and at room temperature, except for the transferred membranes. Membrane transfer was performed at 4˚C with a cold pack and prechilled buffer. Band detection was performed using the enhanced chemiluminescence detection kit (Amersham Pharmacia Biotech GmbH, Freiburg, Germany).
Data Analysis
For confirming the expression of 5-HT1A receptors, the detected bands were quantified densitometrically using Molecular Analyst ver. 1.4.1 (Bio-Rad). The numbers of 5-HT and TPH-positive cells in the dorsal raphe were counted hemilaterally under a light microscope (Olympus, Tokyo, Japan). The data were analyzed with one-way analysis of variance followed by the Duncan post hoc test. All values are expressed as the mean±standard error of the mean. P-value <0.05 was considered significant.
RESULTS
Effect of Treadmill Exercise on Depressive State
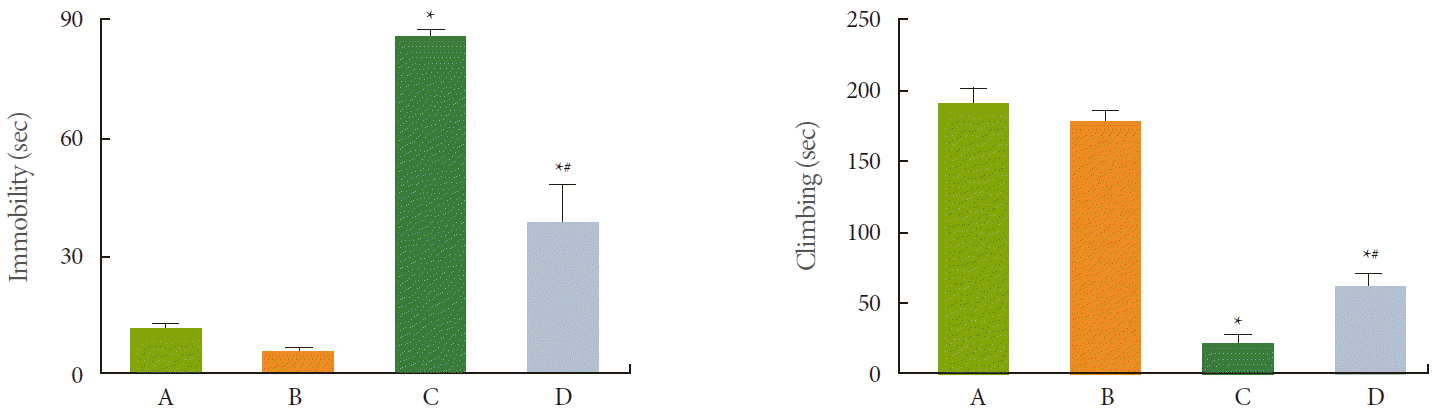
The results of the forced swimming test are presented in Fig. 1. The average time of immobility in the control, control and exercise, stress-induced, and stress-induced and exercise groups were 9.28±2.00, 4.92±0.98, 78.81±10.67, and 43.21±10.56 seconds, respectively. The average time of climbing in the groups was 189.52±13.00, 180.29±8.90, 20.00±4.07, and 62.08±10.70 seconds, respectively. Induction of stress increased the immobility time and decreased the climbing time (P<0.05). In contrast, treadmill exercise decreased the immobility time and increased the climbing time in the stress-induced rats (P<0.05).
Effect of Treadmill Exercise on 5-HT Expression in the Dorsal Raphe
Photomicrographs of 5-HT-positive cells in the dorsal raphe are shown in Fig. 2. The number of 5-HT-positive cells in the control, control and exercise, stress-induced, and stress-induced and exercise group were 299.7±8.42, 277.70±5.14, 170.10± 5.49, and 210.30±6.00, respectively. Induction of stress reduced the expression of 5-HT in the dorsal raphe (P<0.05). In contrast, treadmill exercise increased 5-HT expression in the stress-induced rats (P<0.05).
Effect of Treadmill Exercise on TPH Expression in the Dorsal Raphe
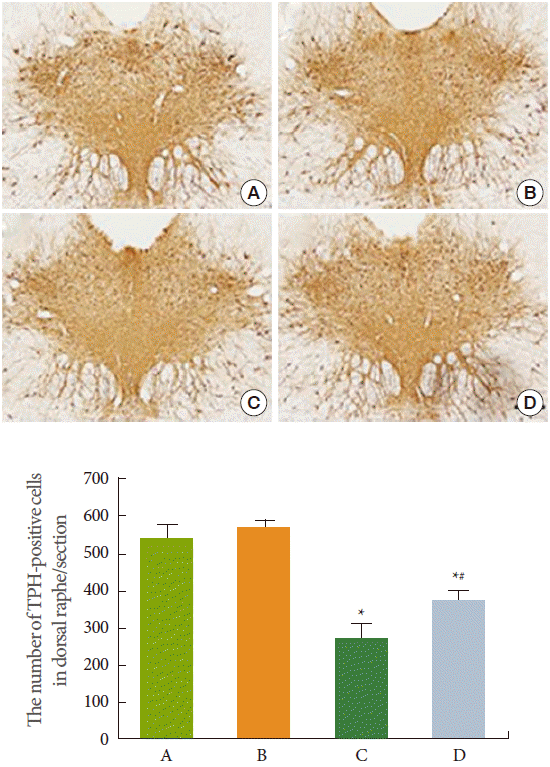
Photomicrographs of TPH-positive cells in the dorsal raphe are shown in Fig. 3. The number of TPH-positive cells in the control, control and exercise, stress-induced, and stress-induced and exercise group were 551.37±45.35, 568.25±37.94, 270.08±36.14, and 381.12±19.10, respectively. Induction of stress reduced the expression of TPH in the dorsal raphe (P<0.05). In contrast, treadmill exercise increased TPH expression in the stress-induced rats (P<0.05).
Effect of Treadmill Exercise on 5-HT1A Receptor Expression in the Dorsal Raphe
5-HT1A receptor expression is shown in Fig. 4. When the level of 5-HT1A in the control group was set at 1.00, the levels of 5-HT1A in the control and exercise, stress-induced, and stressinduced and exercise group were 0.92±0.02, 0.67±0.05, and 0.87±0.06, respectively. Induction of stress reduced the expression of 5-HT1A in the dorsal raphe (P<0.05). In contrast, treadmill exercise enhanced 5-HT1A expression in the stress-induced rats (P<0.05).
DISCUSSION
Exercise is effective for reducing the symptoms of depression and anxiety [21]. A decrease in climbing time and an increase in immobility time in the forced swimming test indicate depressive state in the tested animals [4]. In the present study, the climbing time decreased and the immobility time increased in the stress-induced rats. Stress-induced depression in the forced swimming test was alleviated by treadmill exercise (Fig. 1). These results demonstrate that treadmill exercise exerted an antidepressant-like effect.
The 5-HT in the dorsal and median raphe nuclei ramifies to most of the brain regions, and 5-HT is implicated in emotions, anxiety, and depression [23]. TPH regulates 5-HT activity, and is used as an indicator of serotonin synthesis [24]. Increase in 5-HT synthesis and TPH expression caused by treadmill exercise is well documented [11,20]. Increment of serotonin synthesis and activity might be involved in the antidepressant effect of exercise in the elderly [25]. Baek et al. [20] showed that treadmill exercise increased 5-HT and TPH expressions and ameliorated depression symptoms. In the present study, 5-HT and TPH expressions in the dorsal raphe were suppressed by exposure to stress, and treadmill exercise enhanced 5-HT and TPH expressions in the stress-exposed rats (Figs. 2,3). These results suggest that treadmill exercise ameliorated stress-induced depression through enhancing serotonin synthesis.
Altered transcription of the 5-HT1A receptors affects the serotonin system and leads to a predisposition to depression. The desensitization of 5-HT1A autoreceptors is implicated in the treatment by antidepressants [13]. Savitz et al. [14] raised the possibility that 5-HT1A receptor dysfunction represents a potential mechanism underpinning major depressive disorder and other stress-related disorders. Greenwood et al. [26] demonstrated that wheel running increased 5-HT1A mRNA levels in the dorsal raphe nuclei. In the present study, expression of 5-HT1A receptors in the dorsal raphe was reduced by exposure to stress, and treadmill exercise increased the expression of 5-HT1A receptors in the stress-exposed rats (Fig. 4). These results suggest that treadmill exercise up-regulated 5-HT1A receptors under stressful conditions.
Overactive bladder symptoms are quite bothersome, especially for women, and they increase depression [27]. Ko et al. [28] suggested that physical exercise is a valuable tool for remarkable improvement of overactive bladder symptoms. The present results demonstrate that treadmill exercise enhances 5-HT synthesis through the up-regulation of 5-HT1A receptors, and improves stress-induced depressive symptoms. Physical inactivity, diabetes, hypertension, dyslipidemia, smoking, and obesity are associated with imbalance in oxidative stress, leading to endothelial dysfunction. Such endothelial dysfunction is present in cases of erectile dysfunction [29]. Activation or upregulation of receptors is strongly implicated in the therapeutic strategy for many diseases [30,31]. Leoni et al. [29] suggested the role of physical exercise in the modulation of endothelial function and its implications on erectile function.
Depression aggravates symptoms of an overactive bladder [7] and patients taking antidepressants complain of sexual dysfunction [22]. In the present study, treadmill exercise improved depression symptoms through enhancing 5-HT1A receptor expression. The present results suggest that treadmill exercise might be helpful for the alleviation of overactive bladder symptoms and might improve sexual function.