INTRODUCTION
Serotonin (5-hydroxytryptamine, 5-HT) is a monoamine neurotransmitter that acts via membrane receptors in the central nervous system, the peripheral nervous system, and nonneuronal cells in blood platelets, the gastrointestinal tract, the endocrine system, the sensory system, and the cardiovascular system [1]. To date, 7 families of 5-HT receptors (5-HT1 through 5-HT7) have been identified based on sequence homologies and effector pathways [1]. 5-HT receptors, with the exception of the 5-HT3 class, are G-protein-coupled receptors, and are classified into 13 distinct subtypes: 5-HT1A, 5-HT1B, 5-HT1D, 5-ht1e, 5-HT1F, 5-HT2A, 5-HT2B, 5-HT2C, 5-HT4, 5-ht5a, 5-ht5b, 5-HT6, and 5-HT7 [2]. 5-HT3 receptors belong to the ligandgated ion channel receptor superfamily, and are divided into 5 subcategories: 5-HT3A, 5-HT3B, 5-HT3C, 5-HT3D, and 5-HT3E [2]. 5-HT receptors have been implicated in the central and peripheral regulation of lower urinary tract function [3,4]; however, the physiological role of each 5-HT subtype has been difficult to investigate because of the lack of receptor subtype-specific agonists and antagonists.
Peripherally, 5-HT has been implicated in the modulation of detrusor contraction in mammals, including humans [3]. In resting preparations of human isolated bladder strips, 5-HT-induced contraction was found to be directly and partly mediated by the 5-HT2 receptor subtype located on the effector cells because the effect was inhibited by ketanserin (a 5-HT2 antagonist) but not by tetrodotoxin [5]. Meanwhile, in the electrical field stimulation (EFS)-evoked contraction of human urinary bladder strips, 5-HT-potentiation of the detrusor contraction has been shown to be mediated via 5-HT receptor subtypes, with the exception of 5-HT1, 5-HT2, and 5-HT3, because methysergide (a 5-HT1/5-HT2 antagonist), ketanserin, and ondansetron (a 5-HT3 antagonist) had no effect on the potentiation [6]. This indirect potentiation of detrusor contraction is thought to be mediated by facilitating acetylcholine (ACh) release from parasympathetic terminals via 5-HT4 and 5-HT7 receptors [7,8]. Thus, in the human bladder, in addition to major patterns of mediation via muscarinic receptors, detrusor contractility is modulated by complex mechanisms, partly involving 5HT receptor subtypes.
A recent study using real-time polymerase chain reaction (PCR) and immunohistochemical techniques showed that the expression of 5-HT2B receptor mRNA was markedly greater than that of other 5-HT subtypes in the human detrusor and that the receptors were distributed throughout the muscle layer [9]. These results suggest the possibility that this receptor subtype plays an important role in the modulation of urinary bladder function. In addition, 5-HT2A mRNA was also expressed in the human detrusor with a low expression level, corresponding to one-seventh of that of 5-HT2B mRNA [9].
The aim of this study was to determine the pharmacological significance of specific 5-HT2 subtypes (i.e., 5-HT2A, 5HT2B, and 5-HT2C) in the potentiation of EFS-induced contraction of human isolated urinary bladder strips. The agonist used was α-methyl-5-hydroxytryptamine (α-Me-5-HT), which shows selectivity for 5-HT2B, 5-HT2C, and 5-HT2A in order, with pKi values of 8.4, 7.3, and 6.1, respectively [10]. To block 5-HT2 receptor transmission, ketanserin and naftopidil were used to determine the receptor subtype predominantly involved in the potentiation of detrusor contraction, as these drugs have a fairly high binding affinity for 5-HT2A/2C receptors (5-HT2A >5-HT2C) and 5-HT2A/2B receptors (5-HT2B >5-HT2A), respectively [11,12]. These drugs, which have been shown to ameliorate micturition disturbances in human patients, were selected to explore their possible further clinical benefits as well as to elucidate the basic scientific questions addressed in this study [13,14].
MATERIALS AND METHODS
Isolated Human Urinary Bladder
The present study was conducted in accordance with the European Council Directive 2006/17/CE regarding technical requirements for the donation, procurement, and testing of human tissues and cells.
Human tissues were obtained from 6 male patients with a mean age of 63±3 years (range, 55–74 years). All patients (negative for human immunodeficiency virus 1–2, human T-lymphotropic virus 1–2, and hepatitis B and C) had undergone a cystoprostatectomy for bladder cancer. The tissues were donated after obtaining the patients’ written informed consent. Clinical information about the patients is presented in Table 1.
Experimental Protocol
Each sample of bladder tissue was inspected by direct visual assessment to confirm that it did not include any parts of the malignancy. The detrusor specimen was cleaned of fat and connective tissues, the urothelium removed, and remaining detrusor muscle cut into several strips (length, 9–10 mm; width, 3–4 mm), which were immersed in 5-mL organ baths (EMKA Technologies, Paris, France) containing Krebs buffer (composition in mM: NaCl, 114; KCl, 4.7; CaCl2, 2.5; MgSO4, 1.2; KH2PO4, 1.2; NaHCO3, 25; and glucose, 11.7; pH 7.4, incubated under 95% O2 and 5% CO2 at 37˚C) and mounted under 1 g (10 mN). The Krebs buffer also contained prazosin (1µM) in order to block α1-adrenoceptors. Contractile responses were measured using isometric tension transducers (it-1, EMKA Technologies), and were recorded using Chart software version 7.3.7 and the PowerLabs 8/30 data acquisition system (AD Instruments Pty Ltd., Castle Hill, Australia), running on a personal computer. EFS was performed through platinum electrodes connected to constant current asynchronous stimulators (Model stm-b01, EMKA Technologies). The tissue samples were allowed to equilibrate for at least 60 minutes, during which time contraction with 80mM KCl was performed on each strip. After a 30-minute washout period, tissues were stimulated by EFS for approximately 30–45 minutes, until a stable plateau of contractile responses was obtained, using the following parameters: constant current, 800 mA; frequency, 10 Hz; train duration, 0.1 ms; 5 seconds of train pulses, every 62.5 seconds.
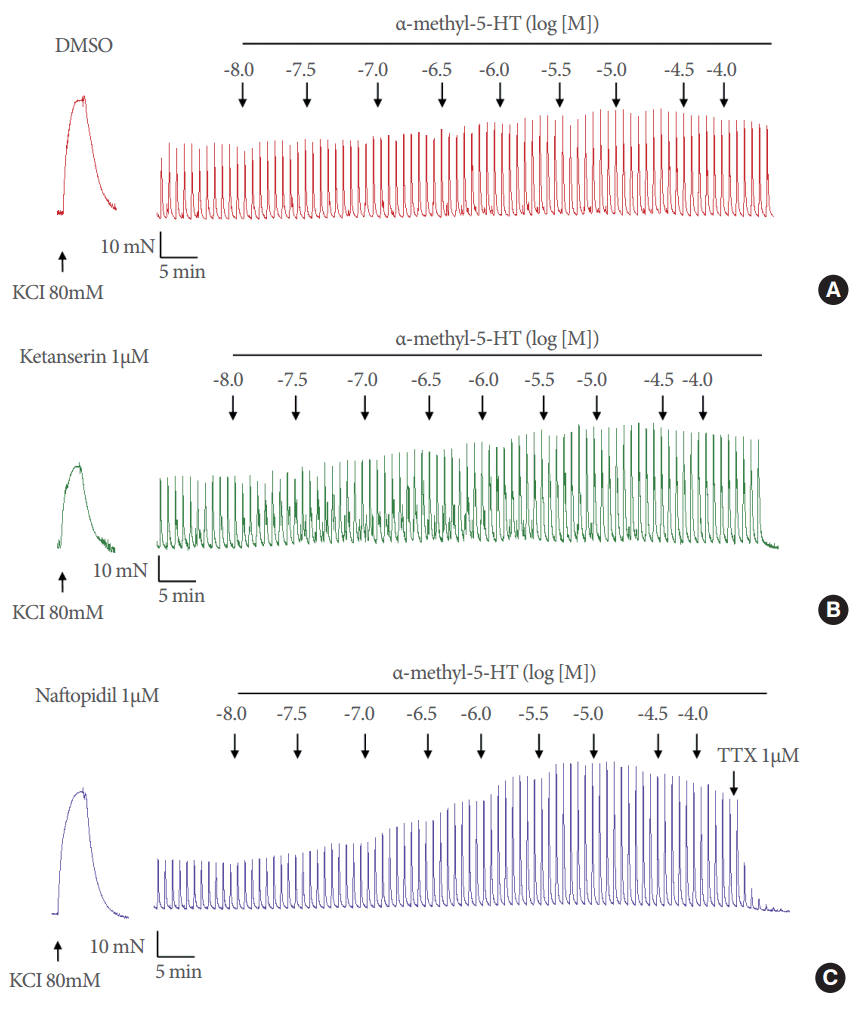
Tissues were stimulated by EFS for 2 serial experiments. In the first experiment, a cumulative concentration-response curve (CRC) for 5-HT or α-Me-5-HT was constructed, in the range of 1nM to 100µM (in half-log increments). The corresponding volume of vehicle was added to strips in the control group. In the second experiment, the cumulative CRC for α-Me-5-HT, in the range of 10nM to 100µM (in half-log increments) was constructed in the presence of ketanserin (0.3, 1, and 3µM), naftopidil (0.3, 1, and 3µM) or their common vehicle, diluted dimethyl sulfoxide (DMSO; 0.4% in distilled water) by incubation for 30 minutes before the first addition of α-Me-5-HT. At the end of both experiments, the nature of the neurogenic contractions was confirmed in at least 2 strips per patient using tetrodotoxin (1µM).
Drugs
5-HT hydrochloride, α-Me-5-HT maleate, ketanserin tartrate, and tetrodotoxin were purchased from Tocris (Bristol, UK). Naftopidil was synthesized at the Asahi Kasei Pharma Corporation (Tokyo, Japan). Prazosin hydrochloride was obtained from Sigma-Aldrich (St-Quentin-Fallavier, France). Stock solutions of naftopidil and ketanserin were prepared on each day of experimentation at a concentration of 0.01M in DMSO. Further dilution was carried out using a mixture of distilled water and DMSO.
Statistical analyses
Values are expressed as mean±standard error of the mean. Responses to agonist concentrations are expressed as the difference between the basal value of contraction induced by EFS (calculated at the end of the stabilization period) and the maximal amplitude of contraction recorded following the addition of each agonist concentration into the organ bath. This delta value (in grams) was normalized against the amplitude of contractions induced by 80mM KCl (in grams), and expressed in terms of percentage of the overall contraction induced by KCl. Fitting and statistical analysis of the results were performed using GraphPad Prism version 6.0 (GraphPad Software Inc., San Diego, CA, USA). The following parameters were determined: the maximal effect induced by the agonist (Emax) and the negative logarithm of the concentration required for a half-maximal response to an agonist (pEC50). The effects of KCl and the amplitude of basal contraction in response to EFS were compared using 1-way analysis of variance. Each CRC for α-Me-5-HT in the presence of ketanserin and naftopidil was compared to the corresponding vehicle CRC using the F-test. A global comparison of the curves was first established. When statistical significance was detected, successive comparisons of Emax and pEC50 values were performed in order to determine which parameter was different. Differences were considered to be statistically significant when the null hypothesis could be rejected at a risk (α) of <0.05.
RESULTS
Potentiation by 5-HT and α-Me-5-HT in EFS-Induced Detrusor Contraction
The tonic and the phasic contractions induced by 80mM KCl and EFS, respectively, among the 5-HT, α-Me-5-HT, and vehicle groups were shown to have no statistically significant differences, and were homogeneous across the groups.
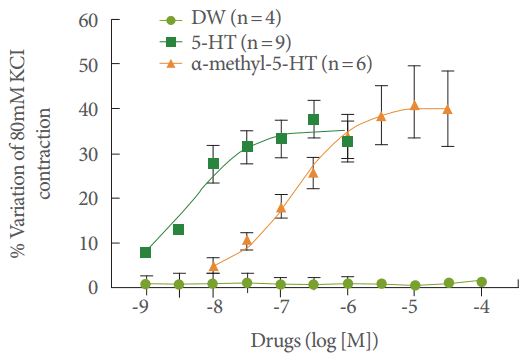
Both agonists (5-HT and α-Me-5-HT) potentiated EFS-induced contractions to a similar extent, with a maximal effect reaching 37.6% ±4.2% (n =9 strips) and 38.6% ±6.6% (n =6 strips), while their pEC50 values were different (8.3 ±0.2 and 6.8 ±0.2, respectively) (Fig. 1). The vehicle had no effect on EFS-evoked contractions (n=4 strips). At the end of the experiments, 1µM tetrodotoxin almost eliminated the EFS-induced contractions of human isolated urinary bladder (82%–98%, n=4 strips), confirming their neurogenic origin. After this set of experiments, α-Me-5-HT was used as a potentiating substance in the next experiment, as it demonstrated an acceptable potentiating response and has been described as having greater selectivity for the 5-HT2 receptor.
Effect of Ketanserin or Naftopidil on Potentiation by α-Me-5-HT in EFS-Induced Detrusor Contraction
In a series of experiments, the bladders of 2 patients were exposed to Bacillus Calmette-Guérin (BCG) therapy before cystectomy (Table 1). A comparison of bladders that did and did not undergo BCG therapy in the CRC for α-Me-5-HT of EFS-induced detrusor contraction was conducted, showing that this treatment did not change the CRC to a significant extent (P=0.066).
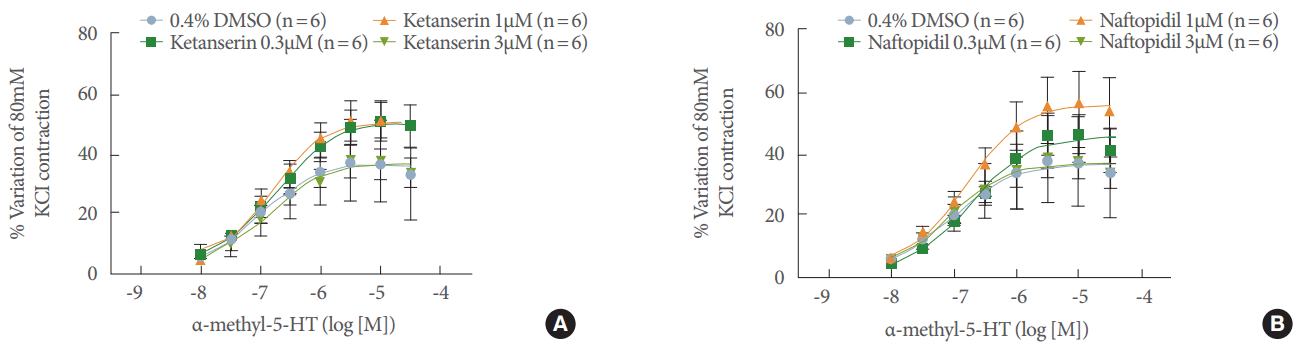
No antagonistic effects on the CRC for α-Me-5-HT were found to result from ketanserin or naftopidil treatment at any concentration (Table 2). However, the maximal effect of α-Me-5-HT was significantly higher in the presence of ketanserin (0.3 and 1µM), with a mean maximal effect of 36.4%±2.2% in the vehicle group, and 50.9% ±2.8% and 51.2% ±3.0% in the 0.3 and 1µM ketanserin groups (P<0.0001 for each), respectively, and in the presence of naftopidil (1µM) with a mean maximal effect of 55.7%±4.1% (P<0.0001) (Table 2 and Figs. 2, 3). A slight additional potentiating effect was observed with 0.3µM of naftopidil, but this effect did not reach significance. At the highest concentration of 3µM, neither ketanserin nor naftopidil had an inhibitory effect on the potentiating activity of α-Me-5-HT. pEC50 values were not significantly modified in the presence of either drug.
DISCUSSION
This study showed that 5-HT and α-Me-5-HT potentiated the EFS-induced contraction of human isolated urinary bladder strips, exerting a similar efficacy, and that ketanserin (0.3 and 1µM) and naftopidil (1µM) further enhanced the potentiating effect of α-Me-5-HT on detrusor contraction, instead of suppressing the effect. The enhancement was produced by a drug concentration close to that reached in plasma concentration when each drug is given at clinically relevant dose, and these results can therefore be suitably translated into bedside practice [15,16]. The contractile responses were mostly neurally mediated, but did not occur owing to direct stimulation of the muscle cells, because the reproducible twitch contractions induced by EFS were eliminated by tetrodotoxin.
The potentiating effect of 5-HT on the EFS-evoked contraction of human isolated bladder strips in this study is comparable to that observed in a previous study by Corsi et al. [6]. Meanwhile, in a study by the same group, α-Me-5-HT behaved as a partial agonist relative to 5-HT in the potentiation of EFS-induced contractions, whereas in our study, it exerted the effect of a full agonist. The cause of this discrepancy is unknown, but may be partly attributable to response differences among individuals, differences in EFS frequency, the reference KCl concentration, the composition of the Krebs solution, or a combination thereof. We added prazosin to the Krebs solution to isolate the effect of facilitation via α1-adrenoceptors on ACh release from the prejunctional parasympathetic nerve terminals and a stimulating effect on postjunctional α1-adrenoceptors in the detrusor [17]. Thus, the increased efficacy of α-Me-5-HT in the present result in comparison with the previous study [6] is not likely to be due to α1-adrenergic mediation.
A recent study using real-time PCR and immunohistochemical techniques revealed that at least 5-HT2A, 5-HT2B, 5-HT3A, 5-HT4, and 5-HT7 were expressed in the urothelium, mucosa, and detrusor, and that 5-HT2B receptors were most common in these layers of the bladder [9]. Taken together with the present result showing that the potentiating effect of α-Me-5-HT was equivalent to that of 5-HT, it may be presumed that 5-HT2B receptors play a major role in the regulation of urinary bladder function. However, 5-HT2 receptor subtypes are not involved in neurally-evoked detrusor contraction because neither ketanserin nor naftopidil suppressed α-Me-5-HT potentiation. Similarly, in a previous study [6], ketanserin at a 1µM concentration did not cause a rightward displacement of the CRC for 5-HT (up to 1µM) in the EFS-induced contraction of human isolated urinary bladder strips.
The contractility or tonus of the detrusor can be locally mediated through multiple complex mechanisms, including a variety of receptors. In the human bladder, the activation of 5-HT4 receptors or 5-HT7 receptors in prejunctional parasympathetic nerve terminals facilitates ACh release [7,8,18], thereby potentiating detrusor contraction, whereas the stimulation of 5-HT1A receptors in cholinergic nerve terminals inhibits its release [8], thereby suppressing contractility. Previously, it has been suggested that α-Me-5-HT interacts with 5-HT receptor subtypes other than 5-HT2, such as 5-HT1A and most probably 5-HT7 [19-21]. Thus, α-Me-5-HT potentiation may be activated via 5-HT7 receptors, although the inhibitory modulation of ACh release by stimulating 5-HT1A receptors is likely, in part, to simultaneously counteract the potentiation. Moreover, α-Me-5-HT has been shown to stimulate β2-receptors [22], which may decrease the detrusor tonus and thus further complicate the interpretation of the present results. It is unknown whether α-Me-5-HT interacts with 5-HT4 receptors.
In this study, neither ketanserin nor naftopidil at any concentration inhibited the α-Me-5-HT potentiation of EFS-induced contraction of human isolated urinary bladder strips. Instead, each drug at low concentrations enhanced the potentiation of the detrusor contraction; ketanserin at 0.3µM and 1µM and naftopidil at 1µM produced maximal potentiation of contractility. What is the underlying mechanism for the enhancement caused by ketanserin or naftopidil of the potentiating effect of α-Me-5-HT? Ketanserin and naftopidil have α1-adrenoceptor antagonism as a common pharmacological property in addition to behaving as 5-HT2 antagonists [23,24]. Both drugs have been shown to possess a high selectivity for α1A- and α1D-adrenoceptors, with a similar affinity [25-27]. Nevertheless, it is not likely that α1-adrenoceptor antagonism, in addition to the prazosin included in Krebs solution, at the prejunctional and postjunctional sites of the detrusor contributed to the enhancement of α-Me-5-HT potentiation. Moreover, it is unknown whether ketanserin and naftopidil effectively activate 5-HT4 receptors, 5-HT7 receptors, or both, which could further enhance the α-Me-5-HT potentiation.
However, ketanserin and naftopidil at the highest concentrations (3µM) completely reversed the enhancement induced by lower concentrations, decreasing it to a level equivalent to the vehicle, Naftopidil has been suggested to possess 5-HT1A agonistic properties [24,28]; thus, it is possible that the drug at 3µM inhibited ACh release by stimulating the inhibitory 5-HT1A mechanism in parasympathetic cholinergic terminals, thereby reversing the enhancement. It is unknown whether ketanserin activates the 5-HT1A receptor mechanism. Further studies are necessary to elucidate the complex mechanisms involving these drugs, which exert their effects via multiple receptors.
In the present study, we used bladder tissue samples obtained from patients who had undergone radical cystectomy for bladder cancer, as is common in studies of the human bladder [6-8,18] due to limited availability and ethical reasons. To date, little is known about whether bladder cancer changes the metabolism of 5-HT and the distribution of 5-HT receptor subtypes in the remaining, nontumor part of the bladder; however, increasing evidence has emerged of links between 5-HT and carcinoma [29]. 5-HT exhibits a growth-stimulatory effect and facilitates cell proliferation in aggressive cancers and carcinoids, often through 5-HT1 and 5-HT2 receptors [30]. It has been suggested that 5-HT1B antagonists, for example, may be useful in the treatment of bladder cancer [30]. In addition, the progression of some tumors is accompanied by dysregulation of the pattern of 5-HT receptor expression [29]. Serum 5-HT levels have been suggested as suitable prognostic marker to evaluate urological tumors, such as urothelial carcinoma in the urinary bladder, adenocarcinoma of the prostate, and renal cell carcinoma [31]. Thus, the possible influence of bladder cancer on the 5-HT-associated motility response of the detrusor may need to be considered.
Two patients received intravesical BCG therapy before cystectomy. BCG induces the production of proinflammatory cytokines, including interferon-γ (IFNγ), tumor necrosis factor-α and interleukin-1β [32,33]. Elevation of IFNγ, which is a dimerized soluble cytokine belonging to the type II class of interferons [32], leads to marked induction of indoleamine 2,3-dioxygenase, a tryptophan-degrading enzyme [34]. This further leads to increased degradation of L-tryptophan via the kynurenine pathway, resulting in decreased 5-HT synthesis [35]. Thus, it is possible that BCG treatment affects the contribution of 5-HT to detrusor motility. In this study, however, BCG-treated bladders showed no difference in the α-Me-5-HT CRC of detrusor contraction compared to what was observed in non-BCG bladders, suggesting that the 4-month interval between the last treatment and cystectomy (and the experiment) was long enough for the functional contribution of 5-HT to be restored in the bladder.
In conclusion, 5-HT2 receptors in the human urinary bladder were not found to participate in the neural modulation of detrusor motor function. It is possible that 5-HT2 receptors are involved in bladder sensory transmission in the afferent pathway, in addition to nonneural detrusor motility. Meanwhile, our results regarding ketanserin and naftopidil were favorable, because these drugs led to enhanced detrusor contraction. Ketanserin has been reported to increase the maximum and mean urine flow rates and to decrease urethral pressure in male patients with prostatism [13]. Naftopidil has been clinically used to treat patients with prostatic hypertrophy and has been also reported to increase the maximum urine flow rate and decrease pressure at the maximum urinary flow rate [14]. Overall, our results suggest the possibility that ketanserin and naftopidil, both of which enhance detrusor contractility and reduce urethral resistance, may be useful for treating prostatism/bladder outlet obstruction, especially concurrent with detrusor hypomotility, as manifested in conditions such as underactive bladder.
HIGHLIGHTS
- The potentiation by α-Me-5-HT of neurally-evoked contraction of human detrusor strips was not mediated via 5-HT2 receptor subtypes, and was enhanced by ketanserin or naftopidil.
- Ketanserin and naftopidil may be useful for treating contractile dysfunction of the detrusor such as underactive bladder.