INTRODUCTION
Micturition requires the participation of subtle sensorimotor regulatory mechanisms for the urinary bladder to be efficiently emptied when socially and behaviorally appropriate. At the nerve-control level, micturition cycles of the bladder filling with urine followed by voiding events requires systematic cooperation among afferent, somatic, and efferent nervous pathways, with efferent pathways stemming from both the central and the autonomic nervous system [1]. Similarly, the muscle-controlled component involves contraction of the bladder detrusor, relaxation of the urethra and the external urethral sphincter, and participation of additional pelvic and perineal striated muscles [2,3]. Other striated muscle regions, especially from the perineal and abdominal muscles, have also been reported to exhibit synergistic activation patterns that could facilitate urination [2,4]. Despite these arguments, the concurrent activation of the abdominal musculature during micturition has remained a controversial topic in the physiology of urination that requires further investigation.
Preclinical studies evaluating the activation of abdominal muscles during micturition using electromyographic (EMG) approaches in rats have provided further information about the physiological requirement of abdominal-visceromotor activity for facilitating voiding events in rodents [5,6]. These observations have broadened our understanding of the voiding reflex, and have raised potential voiding dysfunction issues regarding abdominal neurectomy and postoperative residual curarization that can often result in either paralysis of abdominal muscle groups or loss of this reflex activity after spinal cord injury [7-9]. Therefore, it is necessary to critically assess, in well-standardized rodent models, the contribution of the abdominal muscles to efficient voiding contractions.
By integrating the use of open cystometry (CMG) and a multielectrode EMG system in urethane-anesthetized female mice, we determined whether concomitant activation of abdominal muscles with detrusor contractions occurred during voiding. Secondly, we attempted to quantitatively determine the functionality of the abdominal muscles during micturition, and finally, we investigated the contribution of the abdominal musculature to functional and effective voiding events by blocking nicotinic-type acetylcholine receptors. Although the anesthetic plane may have affected the neurological control of the lower urinary tract, the results of this study could be useful for better understanding the physiology of micturition in rodents, and for determining the effectiveness of new treatments aimed at improving micturition.
MATERIALS AND METHODS
All procedures performed in this study were approved by our Institutional Animal Care and Use Committee (IACUC; ID: AUP-0615-0044/IS00001252), and performed in accordance with the guidelines from the National Institutes of Health on the care and use of laboratory animals [10].
Animals
A total of 6 female C57BL6/J mice (24 to 26 weeks old) were tested in this study. Animals were housed in a pathogen-free environment, with 12-hour light/dark cycles, a controlled room temperature of 25ňöC, and ad libitum access to food and water in plastic cages containing corncob bedding.
Surgical Preparation for CMG
General anesthesia was induced after bilateral subcutaneous injections of urethane (1.2 g/kg) in the thoracic area. Although the voiding responses in urethane-anesthetized animals are not comparable to those seen in awake animals, this anesthetic helps to maintain functional responsiveness of the spinal micturition reflex in rodents [11]. After reaching the anesthetic plane (about 45 minutes), a small abdominal incision was carefully performed to expose the urinary bladder, with special care taken to minimize tissue damage and to keep the rectus abdominis muscle as intact as possible (<1-cm incision). A suprapubic catheter (PE-50) was placed into the bladder dome and fixed with a 6-0 surgical suture. The abdomen incision was then sutured (5-0 silk suture) while leaving the skin flaps open. The bladder catheter was connected to a 3-way valve that was used for the intravesical infusion of saline via an infusion pump, and for the simultaneous measurement of bladder pressure using a transbridge amplifier (World Precision Instruments, Sarasota, FL, USA). After verifying the absence of leaks, the urinary bladder was fully emptied before starting each experiment.
Surgical Preparation for EMG
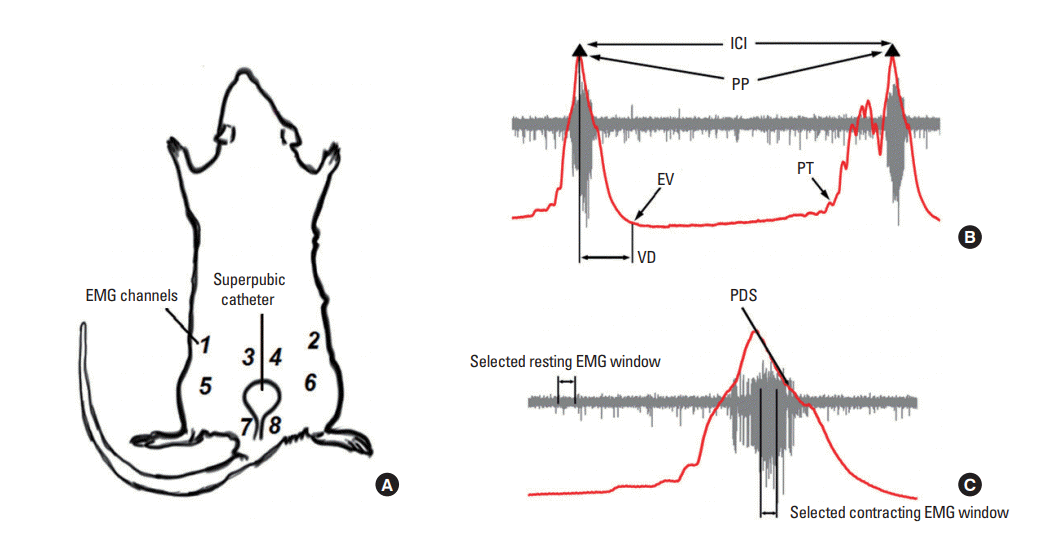
Eight custom-made stainless steel spring electrodes (internal diameter, 0.457 mm) were placed on different muscle surfaces to obtain bilateral EMG recordings from 4 different abdominal muscle domains: the external oblique (EO), internal oblique (IO), and the superior and inferior regions of the rectus abdominis (sRA/iRA) muscles, as shown in Fig. 1A [12]. The placement of all electrodes was verified with a stereomicroscope before initiating the CMG/EMG evaluations. An additional spring electrode was positioned on the chest, in proximity to the heart region, to make electrocardiogram (ECG) recordings and to monitor the vital condition of each animal.
Simultaneous CMG and EMG Data Acquisition
Both EMG and ECG recordings were amplified with a RHD2000 amplifier (Intan Technologies, Los Angeles, CA, USA) at a sampling rate of 20 kHz and gain of 200. Signals for CMG were simultaneously recorded at 5 kHz using an auxiliary input port on the amplifier, where the transbridge amplifier was connected. All electrical and cystometric signals were acquired during constant saline solution (0.9% NaCl) infusion at rate of 20 őľL/min.
Inhibition of Nicotinic Receptors in the Abdominal Surface
After bladder filling with saline to trigger a first voiding, 5 additional voiding events were recorded as described above. Then, without stopping the infusion pump, a solution of pancuronium bromide (Sigma-Aldrich, St. Louis, MO, USA) was carefully applied to the surface of each muscular region where recording was being conducted. Using a micropipette, a total volume of 50 őľL of pancuronium solution per abdominal side was applied on each muscle region. An initial concentration of 25 őľg/mL (the Panc-25 group), followed by 50 őľg/mL pancuronium (the Panc-50 group) was applied successively for partial and complete blockage of muscle activity, respectively. No washouts were performed between drug applications. Five additional voiding events were recorded after each application of the neuromuscular blocking agent. Values were acquired and compared at 3 different stages: intact (only saline infusion), and after the application of 25- and 50-őľg/mL pancuronium. One animal died shortly after the application of 50-őľg/mL pancuronium, so only 1 voiding event was recorded and included in the group analysis. In this case, we suspect that the highly concentrated pancuronium solution perfused into the abdominal cavity, causing rapid cardiac arrest. At the end of the evaluations, the animals were euthanized with CO2 while remaining under deep urethane anesthesia.
Data Analysis
All EMG and CMG data were simultaneously acquired using an Intan interface software and saved for offline signal processing, as described below. Briefly, the CMG signals were downsampled to 62.5 Hz and smoothed with a 15-data-point window length to remove the high-frequency components using MatLab (MathWorks, Natick, MA, USA). Because the animals were in a supine position for electrode placement, the cystometric parameters (Fig. 1B) were calculated from the pressure profile and included the peak pressure for bladder contractions (cm H2O), intercontractile intervals (seconds), pressure threshold (cm H2O), voiding duration (seconds), and pressure drop slope (cm H2O/sec; Fig. 1C). These values were determined in an automated manner using custom-made MatLab scripts. All voiding events were analyzed for the intact conditions and after the application of 25- or 50-őľg/mL pancuronium. The EMG and ECG signals were down-sampled to 5 kHz, and comb-filtered at 60, 120, 180, 240, and 300 Hz to remove powerline noise and harmonics. Thereafter, a second-order Butterworth band-pass filter from 10 to 1,000 Hz was applied. A template subtraction method was employed to remove ECG artifacts in the EMG channels [13], followed by an independent component analysis denoising technique to further secure the elimination of baseline noise and residual ECG artifacts. A threshold of 5 times the standard deviation of the baseline was used to determine both the onset and offset of abdominal muscle activation in the denoised signal [14,15]. A 1-second window was then selected for evaluating both the resting and voiding periods and for calculating the root mean square (RMS) value of each voiding event on each EMG channel (Fig. 1C). The temporal delays between the EMG onset and the CMG pressure peak, from the pressure peak to the EMG offset, and the pressure threshold for EMG onset/offset were also calculated to evaluate simultaneous temporal correlations. Because the EMG amplitudes were not symmetric on either the left or the right flank, and to reduce bias for interpretation, the data were analyzed based on the 4 muscle groups instead of the 8 recording channels.
Statistical Analysis
The 2-tailed paired Student t-test was used to compare the statistical differences in the CMG and EMG parameters before and after the application of each dose of pancuronium. Values are shown as the mean¬Īstandard error of the mean for the voiding events under each condition and animal. The level of statistical significance was set as P<0.05.
RESULTS
EMG Activity of Abdominal Muscles During Voiding
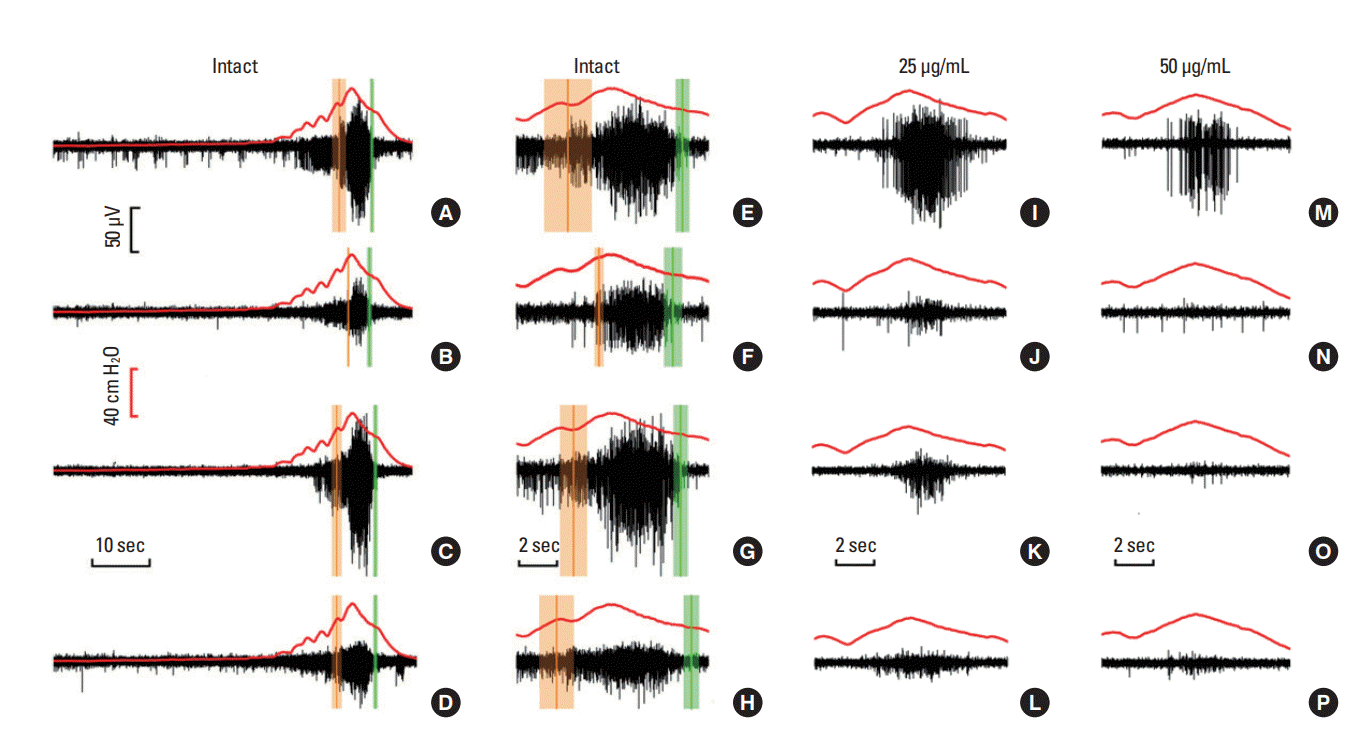
Representative overlaid plots of EMG (black trace) activity and simultaneous CMG activity (red trace) during a voiding event are shown in Fig. 2. The pressure data are presented for a single peak voiding pressure, where high-frequency oscillations were observed after reaching the maximal pressure in all cases. The 4 abdominal muscles were activated concurrently with detrusor contractions during voiding events. Specifically, robust EMG discharges were recorded during micturition events for the EO (Fig. 2A), iRA (Fig. 2B), and IO (Fig. 2C) muscles, while the EMG signals remained low for the sRA (Fig. 2D). Higher timeresolution traces are shown in Fig. 2E‚ÄďH. After the application of 25-őľg/mL pancuronium, both resting and contracting EMG amplitude were attenuated for the EO (Fig. 2I), iRA (Fig. 2J), IO (Fig. 2K), and sRA (Fig. 2L) muscles. Further application of 50-őľg/mL pancuronium eliminated EMG signals in the iRA (Fig. 2N), IO (Fig. 2O), and sRA (Fig. 2P) muscles, with residual activity in the EO muscles (Fig. 2M). Unexpectedly, clear differences in both amplitude and firing patterns were observed in the muscle-region paired electrodes.
Changes in EMG RMS Values
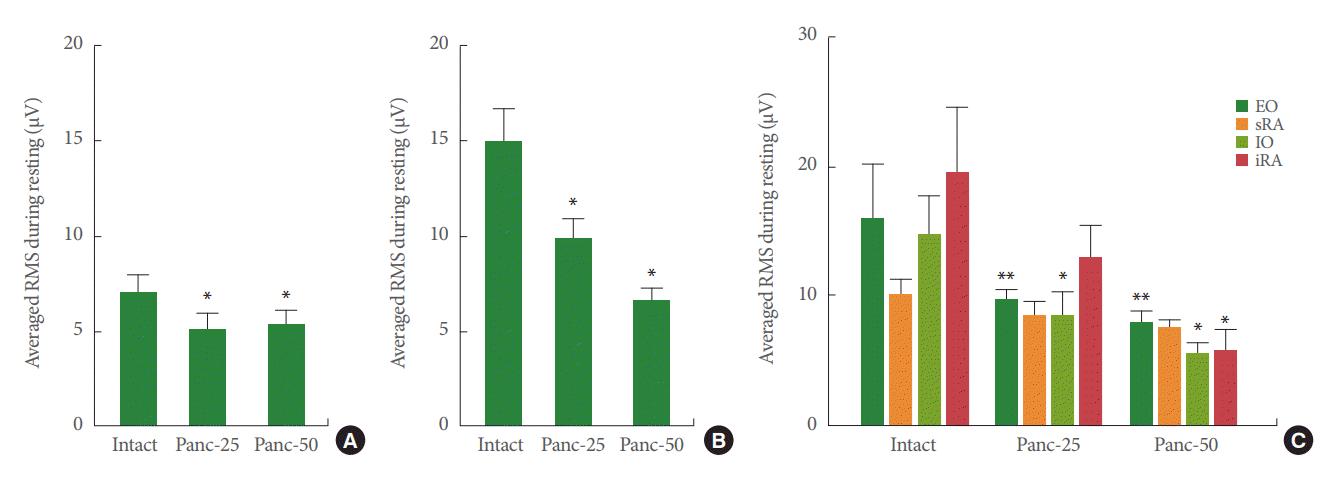
The RMS value in skeletal muscles has been considered to be an indication of signal amplitude, and therefore associated with the strength of motor unit activity during EMG evaluations. Because the automated detection of EMG discharge onset/offset often failed to recognize minor firing events, a 1-second window after reaching the peak pressure value was selected for calculating the voiding RMS. The overall average RMS value for all 4 abdominal muscles was significantly reduced during resting conditions by the application of pancuronium independently of the concentration (Fig. 3A; P<0.05 for both Panc-25 and Panc-50 vs. intact). Moreover, the application of pancuronium on the abdominal surface reduced the average RMS value on all muscles during voiding contractions in a dose-dependent fashion (Fig. 3B; P<0.05 for Panc-25 vs. intact, and P<0.01 for Panc-50 vs. intact). Similarly, in the individual abdominal regions, we found that the RMS during voiding contractions was significantly reduced by the application of pancuronium in the EO, IO, and iRA muscle regions, but no significant changes were observed for the sRA muscle after neuromuscular action blockage (Fig. 3C).
Onset and Offset Times for Abdominal EMG Signals During Voiding
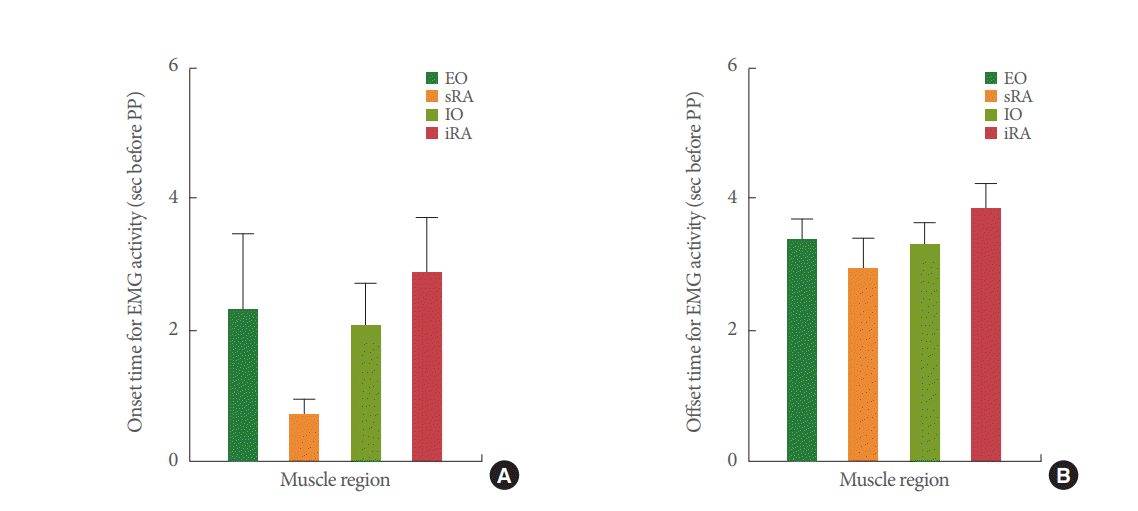
A gradual rise in EMG amplitude during the bladder pressure increase, specifically from the pressure threshold to the peak pressure (pink shadows in Fig. 2), was observed. Automated onset detection analysis showed that the EMG responses of the EO, sRA, IO, and iRA muscles were activated at similar time points before the pressure peak was reached (Fig. 4A). Similarly, the offset times for the EO, sRA, IO, and iRA EMG recordings (green shadows in Fig. 2) were similar for all muscles with respect to the peak pressure (Fig. 4B). Although the IO showed reduced onset values, no significant differences were found in the overall onset and offset time across muscles and conditions (P>0.05). The EMG onset with respect to bladder threshold pressure was calculated as 25.05¬Ī1.35 cm H2O, whereas the offset was 23.22¬Ī0.74 cm H2O. No statistically significant differences were determined for these 2 values.
Changes in CMG Properties After Neuromuscular Blockage
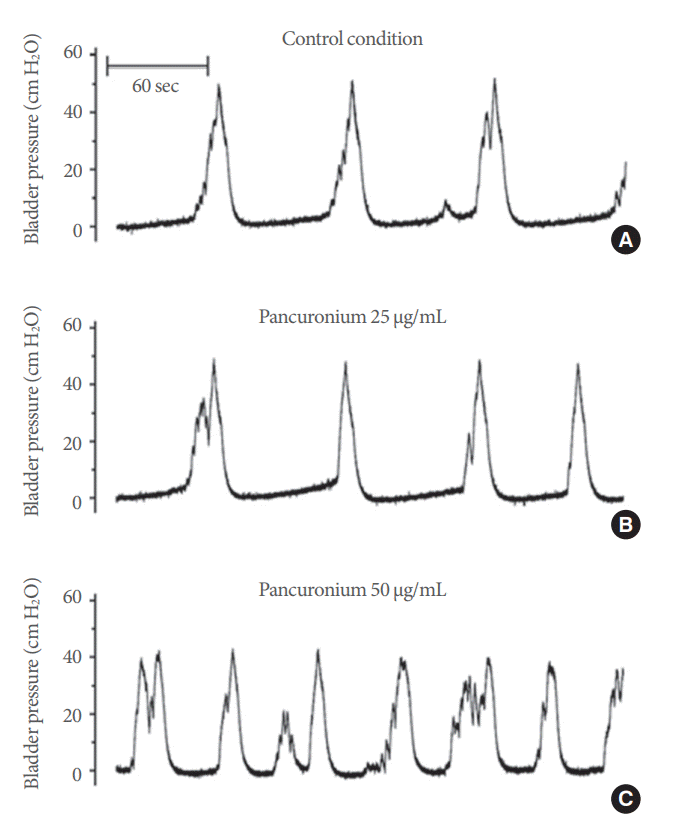
Representative CMG responses from the same mouse are shown during control perfusion (Fig. 5A), and after the application of either 25 őľg/mL (Fig. 5B) or 50-őľg/mL pancuronium to the abdominal muscle (Fig. 5C). Group analysis of the primary cystometric parameters in intact conditions and after the application of 25- or 50-őľg/mL pancuronium are shown in Fig. 6. Values for peak bladder pressure, threshold pressure, and intercontractile intervals were only reduced after the surface application of 50-őľg/mL pancuronium (Fig. 6A, B, and C, respectively). Voiding duration, calculated as the beginning and end of a bladder contractile event, was significantly reduced by either concentration of the nicotinic receptor antagonist (Figs. 5A‚ÄďC and 6D). No significant differences were found for the pressure drop slope in the intact (-3.81¬Ī0.61 cm H2O/sec), Panc-25 (-3.99¬Ī0.56 cm H2O/sec), or the Panc-50 (-3.80¬Ī0.62 cm H2O/sec) groups.
DISCUSSION
Abdominal muscles participate in pelvic expulsive functions, such as defecation and labor [4,9,16,17]. The findings of the present study corroborate that abdominal muscles were also activated during micturition in a female mouse model, and demonstrate that this reflex seems to have exerted a synergistic effect on the efficiency of voiding events. Previous studies have demonstrated that abdominal visceromotor activity was maintained by the pelvic afferent sensory branch, and that activity was intensified by acetic acid through chemical-sensing receptors [5,18]. Our results suggest that abdominal muscles were reflexively activated when the intravesical pressure surpassed approximately 25 cm H2O, with an overall duration of 3 to 5 seconds. As a result, the relatively consistent onset pressure may also suggest that motor neurons innervating the abdominal muscles are recruited through bladder mechanoreceptors when bladder pressure is increased. Previous studies in humans have reported that during voluntary contractions, an initial onset of EO is seen, followed by the pelvic floor and RA muscles, and finally by IO activity [19]. However, our results show varying, but not significantly different, onset and offset times in reference to the pressure peak for the EO, IO, and iRA muscle responses. The difference in the sRA activation period is presumably due to the low EMG amplitude recorded, as shown in Fig. 2B. Although activation can be observed in sRA muscle recordings, the low signal-to-noise ratio greatly impaired the distinguishability of activation periods from resting stages. These results may be considered as further evidence that the abdominal activity during voiding is a reflex event, and different from voluntary contractions, although the physiological differences between human and rodent models need to be considered for an integrative interpretation.
Abdominal muscle straining during voiding in humans is usually considered to be a symptom of bladder outflow obstruction [20,21]. However, clinical observations have failed to demonstrate a consistent relationship between abdominal straining and micturition in healthy subjects or in those with bladder outlet obstruction, suggesting the possibility of nonpathological muscle activity [4,22,23]. The maintenance of abdominal pressure has also been later proven to be an important factor in promoting vesical contraction in humans [24], as well as in animal investigations that confirmed the function of abdominal musculature in improving voiding efficiency [5,6]. For example, it was suggested that contraction of abdominal muscles also contributed to the activation of the pelvic musculature, especially the levator ani muscle, which consequently affected the maintenance of urethral pressure during voiding [4,5]. Similarly, preclinical animal models discovered concurrent discharges of abdominal muscles in both awake and anesthetized female rats, and further investigated the role of sensory pathways in modulating abdominal muscle activity [2]. Based on these studies, a distinct role for the activity of nociceptive abdominal visceromotor responses from that of voiding-associated abdominal responses has been proposed in female rats [6,22], and the authors believed abdominal activity to be important for maintaining high-frequency oscillations as a key feature of voiding patterns in anesthetized rats. Supporting these observations, the recording of maximal intra-abdominal pressure during voiding onset reinforced the necessity of constant abdominal musculature straining for voiding events in awake rats [25].
Altogether, our results and those of the reports discussed above further the idea that abdominal muscles are required for micturition in rodents, as ‚Äėchemical silencing‚Äô with pancuronium significantly affected voiding efficiency. In addition to its supportive role in facilitating urination, a seemingly contradictory function of abdominal straining in maintaining urinary continence has been reported [26,27]. The specific contraction pattern of abdominal muscles, as well as the coactivation of pelvic floor muscles, can contribute to controlling urine flow by, on the contrary, increasing urethral pressure [28,29]. These results have been supported by studies of various therapies involving abdominal muscle training for preventing and treating urinary incontinence [27]. Nonetheless, how the coactivation of 2 muscle groups has different outcomes in reflexive and voluntary activity requires further exploration.
Previous studies evaluating the functional effects of the neuromodulation of pathways innervated by the pelvic nerve on the pelvic musculature have suggested that abdominal muscles play a role in the micturition process [5,18]. Based on bilateral innervation by the pelvic nerve, as well as the bilateral placement of the recording electrodes, a symmetric EMG pattern was expected for each recording region in the mouse abdomen. However, we did not find that to be the case because clear distinctions in amplitude and firing patterns were detected in the muscle-region paired electrodes. We speculate this could have been caused by various factors that are hard to control experimentally, such as differences due to the manual placement of electrode locations and variations in the impedance of the tissue-electrode interface, the intramuscular content of abdominal fat, and cholinergic innervation. Therefore, to reduce bias in data interpretation, our results were analyzed based on 4 muscle groups instead of 8 recording channels. Based on the above considerations, it can be suggested that the multiple-electrode placement used in this study may be an important technique for obtaining more comprehensive information, especially when studying isolated muscular activity in abdominal muscles.
The CMG evaluations during saline infusion suggest that the abdomen plays a significant role in voiding; nonetheless, the targeted blockage of abdominal muscular activity further provided a more specific understanding of the role of abdominal visceromotor activity in facilitating micturition. Our study suggests that local paralysis of the neuromuscular junction is a very useful tool when evaluating abdominal muscle physiology. A parallel approach using botulinum toxin A to induce paralysis of the rectus abdominis muscle resulted in compromised voiding efficiency with decreased voided volume and average urine flow [6]. However, because both the EO and the IO muscles discharge similarly and intensively during voiding, simple blockage of the rectus abdominis is not enough to provide evidence supporting the importance of the full abdomen for micturition.
Pancuronium bromide has been employed in multiple studies as a neuromuscular blocking drug that prevents the activation of acetylcholine receptors [30-32]. However, as pancuronium may also impact the activation of muscarinic receptors in smooth muscle, we decided to apply it onto the abdominal surface instead of intravenously to avoid negative effects on the bladder, heart, and respiratory function [33,34]. The concentrations of the applied pancuronium were selected after preliminary testing in 3 mice that were not included in this report. We found that anesthetized female mice could maintain consistent voiding patterns at a pancuronium concentration of 25 őľg/mL for more than 20 minutes, but after application of a pancuronium solution of 50 őľg/mL, most of the muscular activity in the recorded abdominal regions was eliminated.
In this study, the chemically-induced paralysis of the abdominal musculature reduced the intravesical pressure peak by 18.9%, the intercontractile interval by 47.8%, the pressure threshold by 50.5%, and the voiding duration by 23.2%, suggesting that voiding efficiency was significantly compromised when abdominal muscles did not participate in the micturition process. As the maximal bladder pressure was compromised, we expected a reduced value of the pressure drop slope. However, no significant differences were found for this specific parameter. These results quantitatively support the importance of the reflexive activation of the abdominal musculature for maintaining micturition efficiency in mice. In patients with urine retention, biofeedback of the pelvic floor musculature is often required for maintaining continence and improving voiding efficiency. Indirectly, our results also support the beneficial effects of abdominal muscle training as a urological rehabilitation alternative that needs to be encouraged [35].
It has been considered that the contraction of the abdominal muscles contributes to the increase in intra-abdominal pressure, consequently affecting the intravesical pressure [6]. Unfortunately, intra-abdominal and urethral pressures were not recorded in this study, which somewhat limited the degree to which we could explore in detail the mechanisms of abdominal muscle discharge. Meanwhile, the voided volume was not measured in this study, so compromised voiding efficiency could only be revealed by indirect evidence, such as decreased peak pressure, intercontractile interval, and voiding duration. Direct measurement of the voided volume would provide more details for helping us to interpret the functionality of abdominal muscles in micturition.
Nonvoiding contractions were observed during bladder filling in 2 of the animals that were tested (not shown). Although the female mice used in this experiment were ~6 months old, no age-related detrusor overactivity was expected. Because of this situation, we believe that the increased number of nonvoiding contractions may have been related to the placement of the suprapubic catheter, which perhaps damaged the sensory nervous axons. Although the EMG activity did not show a close correlation with the nonvoiding bladder pressure fluctuations, it is a very interesting observation that an increased baseline RMS value was found in these animals, further suggesting the plausibility of an association of abdominal muscle activation with bladder overactivity. Whether these EMG findings of the abdominal muscles are reproducible in humans, or can be used as a marker for overactive bladder diagnosis and prognosis, remains a topic for future experimental evaluations in healthy patients and those with urological symptoms.
Our results support the presence of a synergistic role for abdominal muscle activation during micturition in anesthetized female mice. The targeted blockage of abdominal muscles, including the EO, IO, sRA, and iRA, proved that they exert a significant function in maintaining voiding efficiency. This study provides an animal model for comprehensively investigating abdominal muscle functions under pathological conditions. Despite the limitations of the present study, our results will advance our understanding of visceromotor activity in the abdominal muscles during voiding.