INTRODUCTION
Chemotherapy is known to induce diverse side effects including damage to the mitochondrial DNA [1]. Doxorubicin (DOX) is a well-known anticancer drug for the treating of solid tumors and hematologic malignancies. DOX is a drug of choice for the patients with breast cancer or prostate cancer. However, DOX is known to cause muscle atrophy and fatigue during therapy [2,3]. Chemotherapy including DOX induces changes of mitochondrial morphology and function in heart, brain, and muscle [4-6]. Although DOX has been used to treat many cancers, its use is limited by its side effects [7,8].
Chemotherapy-induced apoptosis demonstrates diverse apoptosis-related biological candidates, such as increased DNA fragmentation in skeletal muscle [9], activation of several caspases (e.g., 3, 6, and 9) [10,11], cytochrome c release [12], and increased Bax/Bcl-2 ratio [9,10,12]. Mitochondrial DNA damage caused by reactive oxygen species (ROS) results in cellular dysfunction [13]. Chemotherapy enhances ROS production and increases intracellular Ca2+ concentration [14]. And then, these events are associated with the elevated levels of tumor necrosis factor-alpha (TNF-α) and other inflammatory cytokines. The production of inflammatory cytokines, local inflammation, and oxidative stress leads to cellular injury, and eventually causes cell death [15,16]. DOX increases the production of inflammatory cytokines such as TNF-α and reduces muscle contraction [15,17]. This drug also increases free radicals in muscle, in turn, reduces antioxidant level, thereby leading to a loss in muscle mass [18,19]. Throughout the course of chemotherapy, DOX causes fatigue by inducing mitochondrial damage, oxidative stress, and apoptosis [15,19,20].
Exercise regimen has been suggested for the maintaining physical fitness against chemotherapy. Exercise training exerts protective effect on chemotherapy-evoked myopathy in skeletal muscle [20]. Exercise reduces the loss of muscle function by increasing antioxidant enzyme level [17,21]. Continuous exercise improves mitochondrial function, which is impaired by chemotherapy [22].
Fatigue is classified as central fatigue (brain) or peripheral fatigue (muscle) based on the site of occurrence. Central fatigue is associated with impaired exercise performance due to change of neurotransmitters [23]. Serotonin is a neurotransmitter that regulates emotions, depression, lethargy, anxiety, and schizophrenia [24]. Serotonin has been suggested to be associated with the exercise-induced central fatigue [25]. In many studies, neurotransmitters, such as serotonin and dopamine, have been proposed as a causative chemical of physical activity-induced central fatigue [26,27]. In this study, we investigated the effects of treadmill exercise on DOX-induced apoptosis and mitochondrial dysfunction in relation to central fatigue. For this study, we used the rat model of DOX-induced muscle damage.
MATERIALS AND METHODS
Experiment Animals
The procedure of this study followed the guidelines of Korean Academy of Medical Science and the regulations of National Institutes of Health and. We obtained approval from the Kyung Hee University Institutional Animal Care and Use Committee (Seoul, Korea) (KHUASP [SE]-15-086). We divided the Sprague-Dawley male rats (8 weeks in age, 200 g in body weight) randomly into 4 groups: control group, control with exercise group, DOX-treated group, and DOX-treated with exercise group.
DOX-Treated Muscle Damage Model
We obtained DOX from Tokyo Chemical Industry (Tokyo, Japan). To induce the experimental model of muscle damage, DOX (2 mg/kg) dissolved in saline was intraperitoneally injected to the rats, 1 time per week for 4 weeks [28].
Exercise Protocol
The rats in the exercise groups began their running on a treadmill. The running intensity composed of warm up during 5 minutes at 3 m/min, main exercise during 30 minutes at 10 m/min and cool down during 5 minutes at 3 m/min with a 0 inclination. The rats performed treadmill running one time per 1 day and 6 days per week, during 4 consecutive weeks.
Grip Strength and Fatigue Index
Using a grip strength meter (BIO-GS3, Bioseb, Vitrolles, France), muscle strength was determined. After the rat pulled the wire with the paw, we pulled the tail of the rat. The maximum grip strength was recorded as gram until the rat did not release the wire when gently pulling the rat’s back. The test repeated 3 times in the one session, and the mean value was used as the data of maximal grip strength for each rat. We performed 6 sessions with one rat for measurement of muscle strength. The fatigue index was calculated according to the following formula. Fatigue index=(1–[5th measurement of muscle strength+6th measurement]/[1st measurement+2nd measurement])×100.
Measurement of Mitochondrial O2 Respiration
High resolution O2 consumption measurement was conducted by the Oroboros Oxygraph-2 K (Innsbruck, Austria) at 37°C, according to the previous study [28]. After permeabilization and washing the gastrocnemius, it was moved to the chamber with 2 mL of MiR05 respiration buffer (60mM C6H11O7K, 130mM sucrose, 1mM ethyleneglycol tetra-acetic acid (EGTA), 10mM K2HPO4, 3mM MgCl2, 1 mg/mL BSA, and 20mM HEPES, pH 7.4). The protocol consisted of 2mM glutamate with 1mM malate (first), 2mM adenosine diphosphate infusion (ADP) (second), 3mM succinate (third). The completeness of the outer mitochondrial membrane was confirmed to be without increased respiration by addition of cytochrome c. We also confirmed that there is no endogenous substrate for permeabilized myotubes prior to protocol. When there is no added substrate, we washed muscle and measured ADP-stimulated respiration. The O2 consumption rate was presented as picomole/min/mg tissue weight.
Measurement of Mitochondria H2O2 Emission
According to the previous study [28], H2O2 emission was determined by ΔF/min at 37°C during state 4 respiration (10-μg/mL oligomycin) through continually monitoring of the oxidation of Amplex Red (excitation/emission λ=563/587 nm). Mitochondrial H2O2 emission was measured by a Spex Fluormax 4 spectrofluorometer (HORIBA Jobin Yvon, Edison, NJ, USA) at 37°C. The experiment was conducted with buffer Z added to Amplex Red, horseradish peroxidase, and oligomycin, an inhibitor of mitochondrial adenosine triphosphate synthase. The mitochondrial H2O2 emission protocol is as follows: 5mM glutamate and 2mM malate for complex I substrate, 10mM succinate for complex II substrate, and 10mM glycerol-3 phosphate for lipid substrate. The H2O2 emission rate was calculated after subtracting the background value from the reference value (standard curve) using the ΔF/min curve value. The result was presented as picomole/min/mg of tissue weight.
Measurement of Mitochondria Ca2+ Retention Capacity
To assess the openness of the permeability transition pore (PTP), mitochondrial Ca2+ retention capacity was determined, according to the previous study [28]. The mitochondrial Ca2+ retention capacity was analyzed using a Spex Fluormax 4 spectrofluorometer (HORIBA Jobin Yvon). By continuous monitoring of ΔF/min at 37°C during state 4 condition, overlaid traces of Ca2+-induced fluorescence changes by Calcium Green-5N were measured. After background ΔF (1μM Calcium Green-5N, 80μM EGTA, 0.5M glutamate, and 0.2M malate in Buffer Z) was established, then we initiated the reaction by adding Ca2+ pulses (30μM) with wavelengths of excitation and emission at 506 and 532 nm, respectively. Before the PTP opening (i.e., release of Ca2+), mitochondrial Ca2+ retention capacity was presented as picomole/mg of tissue weight.
Tissue Preparation
In order to sacrifice the rats, Zoletil 50 (10 mg/kg; Vibac Laboratories, Carros, France) was intraperitoneally injected into the rats for the anesthetizing. And then, 50mM phosphate-buffered saline (PBS) was transcardially infused. The rats were fixed using 4% paraformaldehyde in 100mM phosphate buffer (pH, 7.4). After the brains were dissected, the brains were post-fixed during overnight and then treated by a 30% sucrose solution. We used a freezing microtome (Leica, Nussloch, Germany) to create sagittal sections of 40 μm thick.
Immunohistochemistry for Tryptophan Hydroxylase and 5-Hydroxytryptamine
We performed immunohistochemistry to calculate the numbers of tryptophan hydroxylase (TPH)-positive and 5-hydroxytryptamine (5-HT)-positive cells in the dorsal raphe, according to the previous study [29]. The sections were treated with PBS for 10 minutes, after that, the sections were washed 3 times by PBS. And then, the sections were treated with 1% H2O2 for 30 minutes, after that the sections were treated to mouse anti-TPH antibody (Oncogene Research Product, Cambridge, UK; 1:500) or to rabbit anti-5-HT antibody (Oncogene Research Product; 1:500) during overnight. The sections were then treated to biotinylated anti-mouse secondary antibody or to biotinylated anti-rabbit secondary antibody (Vector Laboratories, Burlingame, CA, USA) for 1 hour and 30 minutes. After incubation of the sections with avidin-biotin-peroxidase complex (Vector Laboratories) for 1 hour and 30 minutes, immunoreactivity was appeared by treating with a solution composing of 0.05% 3,3’-diaminobenzidine and 0.01% H2O2 in 50mM Tris-buffer (pH, 7.6) for 3 minutes. At room temperature, the slides were airdried during overnight, and Permount (Thermo Fisher Scientific Inc., Waltham, MA, USA) was used to mount.
Western Blot Analysis for Bax, Bcl-2 and Caspases-3
According to the previous study [30], we conducted western blotting. Tissue samples obtained by harvesting of gastrocnemius muscle were lysed using protein lysis buffer. Colorimetric protein assay kit (Bio-Rad, Hercules, CA, USA) was used for the measuring of protein concentration. Protein (20 μg) was separated on sodium dodecyl sulfate-polyacrylamide gels, then it was transferred to a nitrocellulose membrane (Schleicher & Schuell GmbH, Dassel, Germany). For the primary antibodies, we used anti-mouse β-actin antibody (Santa Cruz Biotechnology, Santa Cruz, CA, USA; 1:1,000), anti-mouse Bax antibody (Santa Cruz Biotechnology; 1:1,000), anti-mouse Bcl-2 antibody (Santa Cruz Biotechnology; 1:1,000), and anti-rabbit caspase-3 antibody (Cell signaling Technology, Danvers, MA, USA; 1:1,000). For the secondary antibodies, we used horseradish peroxidase-conjugated anti-mouse antibodies (Santa Cruz Biotechnology; 1:3,000) for β-actin, Bax, Bcl-2, and anti-rabbit antibody (Santa Cruz Biotechnology; 1:5,000) for caspase-3. We calculate the detected bands using enhanced chemiluminescence detection system (Amersham Pharmacia Biotech GmbH, Freiburg, Germany).
Statistical Analysis
The results were statistically analyzed by IBM SPSS Statistics ver. 21.0 (IBM Co., Armonk, NY, USA). For the comparison among the groups, we used 1-way analysis of variance following Tukey post hoc test. Data are presented as the mean±standard error of the mean. Statistical significance was considered when P-value was less than 0.05.
RESULTS
Muscle Strength and Fatigue Index
Fig. 1 shows the results of muscle strength and fatigue index. Muscle strength was decreased by DOX treatment (P<0.05), in contrast, treadmill exercise increased muscle strength in the DOX-treated group (P<0.05). Fatigue index was increased by DOX treatment (P<0.05), in contrast, treadmill exercise decreased fatigue index in the DOX-treated group (P<0.05).
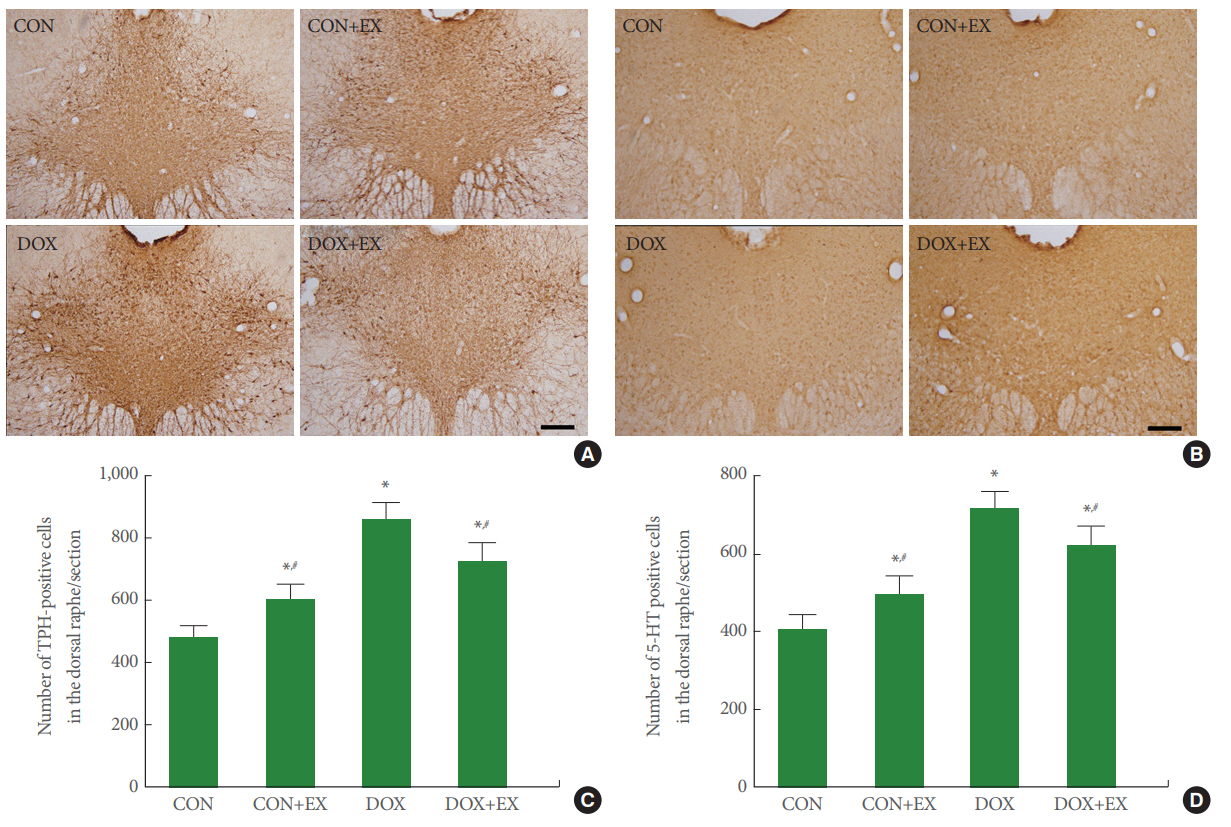
Expressions of TPH and 5-HT in Dorsal Raphe
Fig. 2 shows the numbers of TPH-positive cells and 5-HT-positive cells in the dorsal raphe. Expression of TPH-positive cells in the dorsal raphe was increased by DOX treatment (P<0.05), in contrast, treadmill exercise decreased expression of TPH-positive cells in the DOX-treated group (P<0.05). Expression of 5-HT-positive cells in the dorsal raphe was increased by DOX treatment (P<0.05), in contrast, treadmill exercise decreased expression of 5-HT-positive cells in the DOX-treated group (P<0.05).
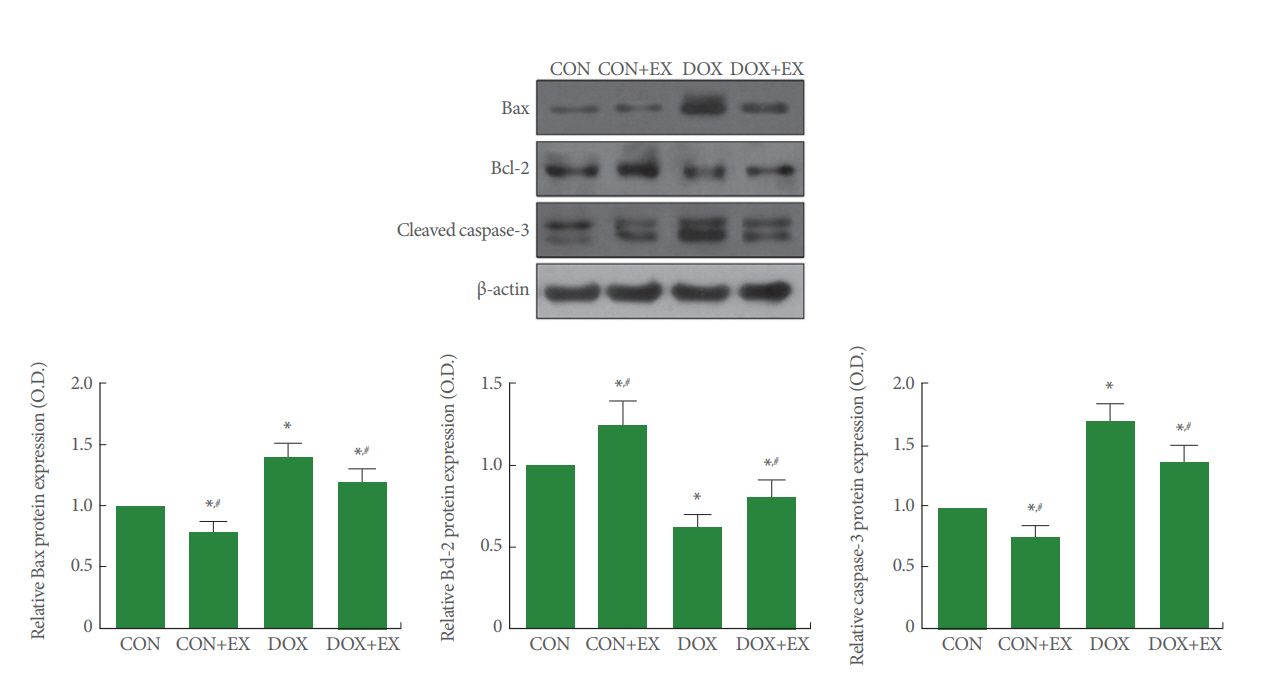
Expressions of Bax, Bcl-2, and Caspases-3 in Gastrocnemius
Fig. 3 shows the expressions of Bax, Bcl-2, and caspases-3 in the gastrocnemius. Bax expression was increase and Bcl-2 expression was decreased by DOX treatment (P<0.05), in contrast, treadmill exercise decreased Bax expression and increased Bcl-2 expression in the DOX-treated group (P<0.05). Caspases-3 expression was increase by DOX treatment (P<0.05), in contrast, treadmill exercise decreased caspases-3 expression in the DOX-treated group (P<0.05).
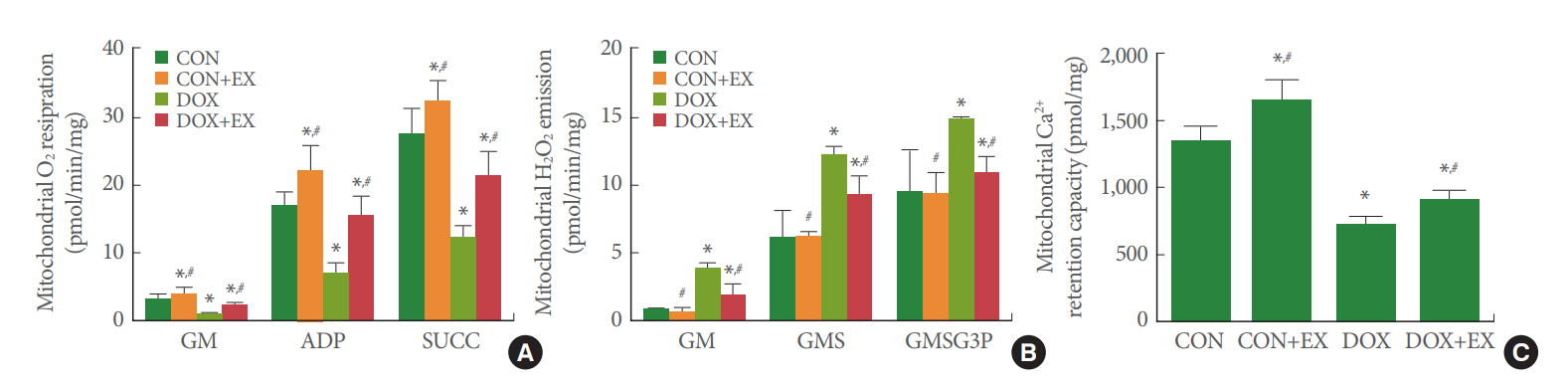
Mitochondrial O2 Respiration in Gastrocnemius
Fig. 4A shows the results of mitochondrial O2 respiration in the gastrocnemius. Mitochondrial O2 respiration under the stage of each factor (glutamate and malate infusion; adenosine diphosphate infusion; succinate infusion) was decreased by DOX treatment (P<0.05). Meanwhile, treadmill exercise increased mitochondrial O2 respiration under the stage of each factor in the DOX-treated group (P<0.05).
Mitochondrial H2O2 Emission in Gastrocnemius
Fig. 4B shows the results of mitochondrial H2O2 emission in the gastrocnemius. Mitochondrial H2O2 emission under the stage of each factor (glutamate and malate infusion; glutamate, malate, and succinate infusion; glutamate, malate, succinate, and glycerol 3-phosphate infusion) was increased by DOX treatment (P<0.05). Meanwhile, treadmill exercise decreased mitochondrial H2O2 emission under the stage of each factor in the DOX-treated group (P<0.05).
Mitochondrial Ca2+ Retention Capacity in the Gastrocnemius
Fig. 4C shows the results of mitochondrial Ca2+ retention capacity in the gastrocnemius. Mitochondrial Ca2+ retention capacity was decreased by DOX treatment (P<0.05). Meanwhile, treadmill exercise increased mitochondrial Ca2+ retention capacity in the DOX-treated group (P<0.05).
DISCUSSION
Anticancer drugs are absolutely necessary treatment for cancer patient, however, the use of these anticancer drugs including DOX is limited by their toxicity. DOX causes side effects such as generation of ROS, stimulation of p53-DNA binding, activation of caspases, DNA cross-linking, and DNA damage [1-3,31].
In this study, enhanced Bax expression and suppressed Bcl-2 expression appeared in the gastrocnemius by DOX treatment, demonstrating that DOX facilitates apoptosis in the muscle. DOX diminishes antioxidant capacity and promotes apoptosis through increasing oxidative stress, and then induces mitochondrial DNA damage [9,11,14]. In this result, Bax expression was suppressed and Bcl-2 expression in the gastrocnemius was enhanced by treadmill exercise, demonstrating that treadmill exercise inhibited DOX-caused apoptotic cell death in the muscle. Suppressing effect of treadmill running on apoptosis is well documented [30,32].
In the present study, O2 respiration and Ca2+ retention capacity in the mitochondria of gastrocnemius were decreased and H2O2 emission in the mitochondria of gastrocnemius was increased by DOX treatment. PTP sensitivity of mitochondria is influenced by ROS production and Ca2+ homeostasis, and mitochondrial function is diminished when apoptosis is exacerbated [14,17]. In the present result, treadmill exercise enhanced O2 respiration and Ca2+ retention capacity in the mitochondria of gastrocnemius and suppressed H2O2 emission in the mitochondria of gastrocnemius, demonstrating that treadmill exercise is effective for the improving mitochondrial function of muscle. Exercise is known to increase mitochondrial plasticity and Ca2+ retention, as a result, exercise alleviates the degree of oxidative stress [9,13].
In the present study, DOX treatment reduced grip strength and increased fatigue index of the gastrocnemius. Increased oxidative stress in the skeletal muscle weakens muscle strength and then causes muscle fatigue [18,19]. In the present result, treadmill exercise increased grip strength and reduced fatigue index, demonstrating that treadmill exercise improves muscle fatigue by reducing DOX-induced muscle weakness. It is known that exercise during chemotherapy can reduce muscle damage caused by DOX [20].
Muscle weakness and muscle fatigue are the common side effects during DOX therapy. Over-expression of TPH and 5-HT in the dorsal raphe is known to be an important factor for central fatigue induction [22,23]. In this study, it was observed that the expressions of TPH and 5-HT in the dorsal raphe were suddenly increased by DOX treatment, suggesting that that DOX therapy induces central fatigue as well as muscle fatigue. 5-HT synthesis and TPH expression in the dorsal raphe are well documented to be enhanced by treadmill exercise [29,33]. In present result, however, treadmill exercise suppressed DOX-induced over-expression of TPH and 5-HT in the dorsal raphe. This suppression on 5-HT synthesis and TPH expression by treadmill running might contribute to the amelioration of DOX-induced central fatigue.
In conclusion, DOX increased apoptosis and decreased mitochondria function in muscle, resulting in muscle weakness and central fatigue. Treadmill exercise suppressed apoptosis and prevented deterioration of mitochondria function in muscle, resulting in alleviation of muscle weakness and central fatigue during DOX therapy.