Exercise Training Attenuates Ovariectomy-Induced Alterations in Skeletal Muscle Remodeling, Apoptotic Signaling, and Atrophy Signaling in Rat Skeletal Muscle
Article information

Abstract
Purpose
The effects of aerobic exercise training on soleus muscle morphology, mitochondria-mediated apoptotic signaling, and atrophy/hypertrophy signaling in ovariectomized rat skeletal muscle were investigated.
Methods
Female Sprague-Dawley rats were divided into control (CON), ovariectomy (OVX), and ovariectomy plus exercise (OVX+EX) groups. After ovarian excision, exercise training was performed using a rat treadmill at 20 m/min, 50 min/day, 5 days/week for 12 weeks. Protein levels of mitochondria-mediated apoptotic signaling and atrophy/hypertrophy signaling in the skeletal muscle (soleus) were examined through western immunoblot analysis.
Results
The number of myocytes and myocyte cross-sectional area (CSA) were increased and the extramyocyte space was decreased in the OVX group compared to those in the CON group. However, aerobic exercise training significantly increased myocyte CSA and decreased extramyocyte space in the OVX+EX group compared to those in the OVX group. The protein levels of proapoptotic signaling and muscle atrophy signaling were significantly increased, whereas the protein levels of muscle hypertrophy signaling were significantly decreased in the OVX group compared to that in the CON group. Aerobic exercise training significantly decreased the protein levels of proapoptotic signaling and increased the protein level of antiapoptotic protein in the OVX+EX group compared to that in the OVX group. Aerobic exercise training significantly increased the protein levels of hypertrophy signaling and decreased protein levels of atrophy signaling in the OVX+EX group compared to those in the OVX group.
Conclusions
Treadmill exercise improved estrogen deficiency-induced impairment in skeletal muscle remodeling, mitochondria-mediated apoptotic signaling, and atrophy/hypertrophy signaling in skeletal muscle.
• HIGHLIGHTS
- Estrogen deficiency induces skeletal muscle remodeling.
- Exercise reverses estrogen deficiency-induced apoptotic signaling.
- Exercise suppresses estrogen deficiency-induced atrophy of skeletal muscles.
INTRODUCTION
The population of women aged 65 years or above is increasing, and their life expectancy has increased to 100 years. Typically, menopause in women occurs between 49 and 52 years of age [1]. Thus, women spend more than one-third of their lives after menopause, which is characterized by low levels of estrogen. Estrogen is a sex steroid hormone that plays a crucial role in human physiology. Therefore, the decline in estrogen levels is related to cardiovascular disease, obesity, type II diabetes [2,3], and osteoporosis [4,5].
In the human body, skeletal muscle is an essential metabolic tissue that accounts for approximately 45% of the whole-body mass. Rolland et al. [6] demonstrated that muscle mass decreased by 0.6% per year after menopause. A previous study reported that decreased estrogen levels influenced skeletal muscles and accelerated the pathogenesis of sarcopenia [7], which is the loss of muscle strength, mass, and physical performance [8]. Akt/mTOR signaling is a regulator of muscle quality, such as muscle strength and physical performance [9]. Protein kinase b (Akt) regulate protein synthesis, which activates the phosphorylation of eukaryotic translation initiation factor 4E binding protein 1 (4E-BP1) and ribosomal protein S6 kinase beta-1 (p70S6K) [10], and inactivates forkhead box O (FOXO) transcription factors [11]. A previous study demonstrated that ovariectomy-induced a decrease in muscle Akt phosphorylation, which was related to a decline in skeletal muscle mass [12]. Apoptosis is induced by 3 pathways: (1) sarcoplasmic reticulum/Ca2+-driven pathway, (2) cytokine/Fas receptor-driven pathway, and (3) mitochondria-mediated pathway [13]. The mitochondria-mediated pathway is the most important regulator of various diseases related to skeletal muscle [14,15]. The Bcl-2 pathway includes antiapoptotic proteins and proapoptotic proteins [13]. The ratio of proapoptotic such as Bcl-2 to antiapoptotic such as Bax proteins manages cellular survival by controlling mitochondrial membrane permeability [13]. Unbalance mitochondrial membrane integrity leads to the release of cytochrome c from the mitochondria into the cytosol, and it activates caspase-3 and caspase-9, which results in DNA fragmentation and cell death [16]. According to previous studies, estrogen has a crucial role in the inhibition of mitochondria-mediated apoptosis in skeletal muscles [17]. However, there are very few studies on the relationship between estrogen and apoptosis.
As a therapeutic intervention, exercise training is well known for its effectiveness in various diseases such as cancer, obesity, and type II diabetes [18-21]. Exercise training is also known to have positive effects on age-related sarcopenia [22,23]. In addition, there are several studies on the effects of exercise on decreased muscle mass induced by estrogen deficiency. Exercise training is considered an important contributor to muscle mass, strength, and quality of life after menopause [24]. Although the importance of exercise training in estrogen deficiency has been established, the effects of exercise training on mitochondriamediated apoptosis and atrophy/hypertrophy signaling in the skeletal muscle have not been elucidated. Therefore, in this study, we hypothesized that moderate aerobic exercise training could ameliorate estrogen deficiency-induced skeletal muscle morphological changes, mitochondria-mediated apoptotic signaling, and atrophy signaling in rat soleus skeletal muscle.
MATERIALS AND METHODS
Experimental Design
Female Sprague-Dawley rats were 7-week-old. Rats were divided into 3 groups (n=6): control (CON), ovariectomy (OVX), and ovariectomy plus exercise (OVX+EX). The sample size was determined using a previous study [25], and the sample size of 6 animals per group was adequate to detect significant results in the current study. Rats were housed under a temperature-controlled room with a 12:12-hour light-dark cycle, and food and water were provided ad libitum. OVX and OVX+EX groups underwent ovarian resection to induce estrogen deficiency at 7 weeks. In the OVX+EX group, exercise training was performed 2 weeks after surgery. After completing treadmill exercise training for 12 weeks, skeletal muscle (soleus) tissues were extracted, and apoptotic signaling and atrophy/hypertrophy signaling were analyzed via western blotting. Histological analysis was used to determine morphological changes after ovariectomy and exercise.
Animals and Ethical Approval
All animal experimental procedures were confirmed with the guidelines of the Korean Academy of Medical Science and the regulations stipulated by the National Institutes of Health (NIH). This study was approved by the Han Nam University Institutional Animal Care and Use Committee (HNU 2018-1).
Ovarian Resection
Rats were anesthetized with ketamine and 2% xylazine, skin areas were shaved, and exposed skin was cleaned with 70% alcohol wipe. An incision of approximately 1 cm was made in the center of the abdomen, and the ovaries were ligated with surgical suture thread. The bilateral ovaries were removed. After ovarian resection, the wound was closed using a synthetic thread, and antibiotics (cefazolin 50 mg/kg) were injected into the muscle to prevent infection. Exercise training was conducted 2 weeks after the surgery.
Western Immunoblot Analysis
The soleus fibers were homogenized using a lysis buffer and centrifuged at 13,000 rpm for 20 minutes. Protein concentration was measured via colorimetric protein assay kit (Bio-Rad, Hercules, CA, USA). Proteins (30 µg) were separated on sodium dodecyl sulfate-polyacrylamide gels and transferred onto a nitrocellulose membrane on ice. The membranes were incubated using 5% skim milk with Tris-buffered saline which is containing 0.1% Tween-20 (TBS-T) and then, incubated overnight at 4°C with the following primary antibodies: GAPDH, Bcl-2, Bax, cytochrome c (Santa Cruz Biotechnology, Santa Cruz, CA, USA), Akt, p-AKT, mTOR, p-mTOR, p70S6k, p-p70S6k, 4EBP1, p-4E-BP1, FOXO3, MuRF1, and Atrogin-1 (Cell Signaling Technology, Danvers, MS, USA). The membranes were incubated for 1 hour with secondary antibodies: horseradish peroxidase (HRP)-conjugated anti-mouse (Santa Cruz Biotechnology) or goat anti-rabbit IgG-heavy and light chain antibody HRP conjugated. After washing 3 times in TBS-T, an enhanced chemiluminescence detection kit (Thermo Fisher Scientific, Waltham, MA, USA) was used to detect the band. Protein bands was expressed using a ChemiDoc (Bio-Rad). All protein levels were calculated with GAPDH as a ratio.
Hematoxylin and Eosin Staining
The paraffin block embedded and sectioned soleus muscle was cut to 5-μm thickness. Hematoxylin and eosin staining was performed at room temperature to localize the geometry, myocyte nuclei, and extramyocyte space. The number of myocytes, myocyte CSA, and extramyocyte space were measured using the Image analysis program (NIH, Bethesda, MD, USA).
RESULTS
Effects of Ovariectomy and Exercise Training on Morphology in Rat Skeletal Muscle
The number of myocytes was significantly lower in the OVX group than in the CON group (P<0.05) (Fig. 1A, B). The myocyte CSA also decreased by 41% in the OVX group compared to that in the CON group (P<0.05) (Fig. 1C). In addition, significantly increased extramyocyte space was confirmed in the OVX myofibers compared to that in the CON myofibers (P<0.05) (Fig. 1D). Conversely, myocyte CSA was significantly increased by 26% (P<0.05) (Fig. 1C). Also, the extramyocyte space was significantly decreased by 26% in the OVX+EX group compared to that in the OVX group (P<0.05) (Fig. 1D).
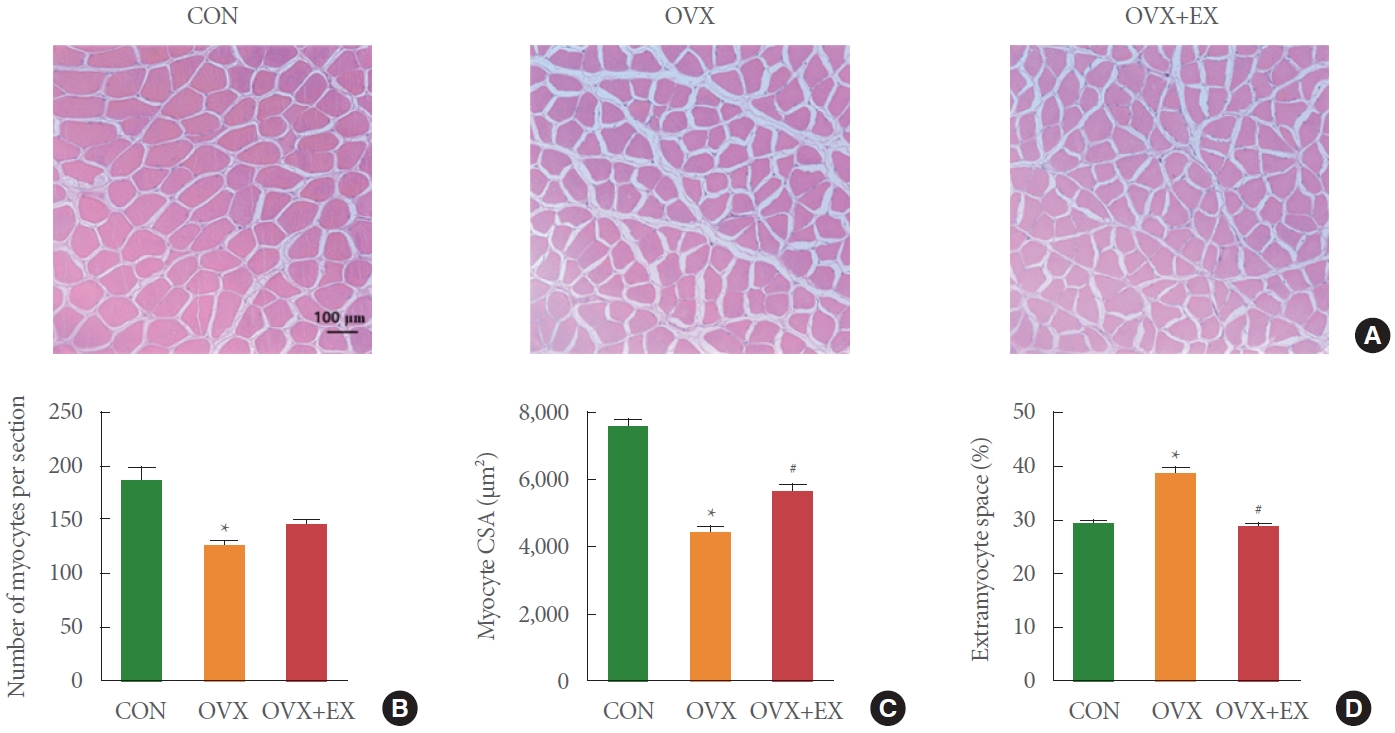
Effects of ovariectomy and exercise training on morphology. (A) Histological finding. (B) Number of myocytes per section. (C) Myocyte CSA. (D) Extramyocyte space. The data are indicated as the mean±standard error of mean. CON, control group; OVX, ovariectomy group; OVX+EX, ovariectomy plus exercise group; CSA, cross-sectional area. *P<0.05 compared with CON. #P<0.05 compared with OVX.
Effects of Ovariectomy and Exercise Training on Mitochondria-Mediated Apoptotic Signaling in Rat Skeletal Muscle
The Bax protein levels in the OVX group were significantly increased by 104% than those in the CON group (P<0.05) (Fig. 2A). In contrast, Bcl-2 protein levels were decreased by 36% in the OVX group than those in the CON group (P<0.05) (Fig. 2B). Taken together, the Bax/Bcl-2 ratio, which is an early stage of mitochondria-mediated apoptosis, was increased by OVX (P <0.05) (Fig. 2C). However, Bcl-2 protein levels were increased by 182% in the OVX+EX group compared with the OVX group (P<0.05) (Fig. 2B), suggesting that aerobic exercise training for 12 weeks protected against OVX-induced increase in early mitochondria-mediated apoptotic signaling.

Effects of ovariectomy and exercise training on mitochondria-mediated apoptotic signaling. (A) Bax. (B) Bcl-2. (C) Bax/Bcl-2 ratio. (D) Cytochrome C. The data are indicated as the mean±standard error of mean. CON, control group; OVX, ovariectomy group; OVX+EX, ovariectomy plus exercise group. *P<0.05 compared with CON. #P<0.05 compared with OVX.
In addition, the protein levels of cytochrome c, which is released by the mitochondrial permeability transition pore (mPTP) opening, also showed similar results. The protein levels of cytochrome c were significantly increased by 105% in the OVX group compared to those in the CON group (P<0.05) (Fig. 2D). However, the protein levels of cytochrome c were significantly lower in the OVX+EX group than in the OVX group (P<0.05) (Fig. 2D).
Effects of Ovariectomy and Exercise Training on Atrophy and Hypertrophy Signaling in Rat Skeletal Muscle
The protein levels of FOXO3 were significantly increased by 62% in the OVX group compared to those in the CON group (P<0.05) (Fig. 3A). Consistently, MuRF1 and Atrogin-1 protein levels were significantly increased by 40% and 88%, respectively, in the OVX group compared with the CON group (P<0.05) (Fig. 3B, C). Conversely, FOXO3, MuRF1, and Atrogin-1 were significantly decreased by 60%, 44%, and 42%, respectively, in the OVX+EX group compared with the OVX group (P<0.05) (Fig. 3A-C), suggesting that 12 weeks of aerobic exercise training attenuated OVX-induced skeletal muscle atrophy.

Effects of ovariectomy and exercise training on atrophy signaling. (A) FOXO3. (B) MurF1. (C) Atrogin-1. The data are indicated as the mean±standard error of mean. CON, control group; OVX, ovariectomy group; OVX+EX, ovariectomy plus exercise group; FOXO3, forkhead box O3; MuRF1, muscle ring finger 1; Atrogin-1, muscle specific f box. *P<0.05 compared with CON. #P<0.05 compared with OVX.
The protein levels of p-Akt/Akt, p-mTOR/mTOR, p-4E-BP1/4E-BP1, and p-p70S6K/p70S6K were significantly decreased by 55%, 37%, 17%, and 39%, respectively, in the OVX group compared with the CON group (P<0.05) (Fig. 4A-D). However, the protein levels of p-Akt/Akt, p-mTOR/mTOR, and p-p70S6K/p70S6K were significantly increased by 128%, 42%, and 25%, respectively, in the OVX+EX group compared with that in the OVX group (P<0.05) (Fig. 4A-C).

Effects of ovariectomy and exercise training on hypertrophy signaling. (A) p-Akt/Ak. (B) p-mTOR/mTOR. (C) p-p70S6K/p70S6K. (D) p-4E-BP1/4E-BP1. The data are indicated as the mean±standard error of mean. CON, control group; OVX, ovariectomy group; OVX+EX, ovariectomy plus exercise group; Akt, protein kinase B; mTOR, mammalian target of rapamycin; p70S6K, ribosomal protein S6 kinase beta-1; 4E-BP1, eukaryotic translation initiation factor 4E binding protein 1. *P<0.05 compared with CON. #P<0.05 compared with OVX.
DISCUSSION
In this study, we demonstrated that aerobic exercise training ameliorated OVX-induced skeletal muscle morphological changes, suggesting that the OVX-induced increase of the connective tissue in the skeletal muscle was reduced and that the decrease in CSA was reversed by aerobic exercise training. Also, we found that aerobic exercise training in OVX rats protected against an OVX-associated increase in mitochondria-mediated apoptotic signaling (e.g., Bax/Bcl-2 ratio and cytochrome c level) in skeletal muscle. In addition, we found that atrophy signaling was decreased, and hypertrophy signaling was increased by aerobic exercise training in ovariectomized rat skeletal muscle. These data support our hypothesis that aerobic exercise training would ameliorate OVX-induced alterations in skeletal muscle morphological changes, mitochondria-mediated apoptotic signaling, and atrophy/hypertrophy signaling in rat skeletal muscle.
Consistent with previous findings [26,27], our study found significant skeletal muscle remodeling, including decreased myocyte number and myocyte CSA, and increased extramyocyte space in the skeletal muscle of ovariectomized rats (Fig. 1), which finally led to the loss of skeletal muscle mass [26]. Many studies have reported the effects of resistance training and hormone replacement therapy as therapeutic interventions for skeletal muscle remodeling [26-28]. However, only a few studies are demonstrating the effect of aerobic exercise, which contributes to increased muscle protein synthesis, on skeletal muscle remodeling in ovariectomized rats. Notably, we observed that aerobic exercise training attenuated estrogen deficiency-induced changes in the myofiber area (CSA) and extramyocyte space. We also observed that aerobic exercise training in OVX rats had no effect on myocyte number, in accordance with previous findings using high-fat diet-induced obesity models [15]. In turn, it seems difficult to expect the effects of aerobic exercise training on skeletal muscle fiber number because skeletal muscles are postmiotic tissues [15]. Therefore, these results suggest that as a therapeutic intervention, aerobic exercise training for 12 weeks exerts beneficial effects on OVX-induced skeletal muscle remodeling.
Mitochondria play an essential role in regulating cell death and survival, also known as mitochondria-mediated apoptosis [29]. We expected that mitochondria-mediated apoptotic signaling could be induced by OVX, and that aerobic exercise training would attenuate mitochondria-mediated apoptotic signaling in the soleus muscles of ovariectomized rats. Indeed, in the current study, the Bax/Bcl-2 ratio was significantly higher in ovariectomized rat skeletal muscle than in nonovariectomy skeletal muscle (Fig. 2C). This increased Bax/Bcl-2 ratio by OVX and led to an elevated release of cytochrome c from the mitochondria (Fig. 2D). Moreover, we demonstrated that aerobic exercise training induced the downregulation of cytochrome c and the Bax/Bcl-2 ratio in ovariectomized rat skeletal muscle (Fig. 2C, D). Taken together, increased cytochrome c and Bax/Bcl-2 ratio were markedly attenuated by 12 weeks of aerobic exercise training, suggesting that aerobic exercise training as a therapeutic intervention protects against OVX-induced apoptotic signaling in skeletal muscles. Further studies are needed to confirm that mPTP opening, caspase-9, and caspase-3 cleavage are key regulators of mitochondria-mediated apoptosis, which ultimately lead to DNA fragmentation and cell death [13].
Skeletal muscle mass depends on the dynamic balance between protein degradation and synthesis. Many studies have clearly confirmed that the Akt/mTOR pathway is important in regulating muscle protein synthesis [30,31], and FOXO3 transcription factor plays a crucial role in muscle atrophy [32], which leads to the expression of MuRF1 and Atrogin-1 [33]. We hypothesized that OVX-induced estrogen deficiency resulted in increased atrophy signaling and decreased hypertrophy signaling in the skeletal muscle. Indeed, the current study demonstrated that muscle atrophy signaling markers such as FOXO3, MuRF1, and Atrogin-1 were increased in the OVX group (Fig. 3). Conversely, in the OVX group, muscle hypertrophy signaling markers such as Akt, mTOR, p70S6K, and 4E-BP1 were decreased (Fig. 4), suggesting that OVX-induced estrogen deficiency results in skeletal muscle atrophy. In addition, we showed that aerobic exercise training attenuated atrophy signaling and recovered hypertrophy signaling in OVX-induced skeletal muscle (Figs. 3, 4). These results suggest that aerobic exercise training protects against OVX-induced skeletal muscle atrophy by activating Akt-mTOR signaling. However, there are only a few studies on aerobic exercise training as a therapeutic intervention for atrophy induced by OVX. Therefore, more studies are needed to verify the effects of aerobic exercise training on the recovery of OVX-induced skeletal muscle atrophy.
Twelve weeks of aerobic exercise training ameliorated OVX-induced changes in skeletal muscle morphology, demonstrating that exercise training ameliorated the OVX-related decrease in myocyte CSA and increase in extramyocyte space. In addition, exercise training in ovariectomized rats protected against OVX-induced increases in mitochondria-mediated apoptotic signaling (e.g., Bax, Bcl-2, cytochrome c) protein levels in the skeletal muscle of ovariectomized rats. Moreover, exercise training attenuated skeletal muscle atrophy signaling (e.g., FOXO3, MuRF1, Atrogin-1) and recovered skeletal muscle hypertrophy signaling (e.g., Akt, mTOR, p70S6K, 4E-BP1) in ovariectomized rats. These findings support the hypothesis that modulation of mitochondria-mediated apoptosis by aerobic exercise training could attenuate OVX-induced skeletal muscle remodeling and thus reduce atrophy of skeletal muscle with estrogen deficiency.
Notes
Fund/Grant Support
This work was supported by the Ministry of Education of the Republic of Korea and the National Research Foundation of Korea (NRF-2016R1A2B4014240, NRF-2018R1A2A3074577, NRF-2019S1A5C2A03082727).
Research Ethics
Animal care and experimental procedures with rats were approved by the Han Nam University Institutional Animal Care and Use Committee (HNU 2018-1).
Conflict of Interest
No potential conflict of interest relevant to this article was reported.
AUTHOR CONTRIBUTION STATEMENT
• Conceptualization: EJC
• Data curation: YC, JK, JHB
• Formal analysis: YC, JK, JHB
• Funding acquisition: HBK
• Methodology: JC, DHP, JHK
• Project administration: HBK
• Visualization: JHY, EP, DYS, SL
• Writing-original draft: EJC
• Writing-review & editing: HBK