ŌĆó HIGHLIGHTS
- Whole-cell patch-clamp recordings were made using SG neurons in spinal cord slices from rats.
- Naftopidil significantly decreased the peak amplitudes of Ad and C fiber-evoked EPSCs, but not prazosin.
- Inhibition of evoked EPSCs by naftopidil may contribute to suppression of the micturition reflex.
INTRODUCTION
Lower urinary tract symptoms (LUTS) are rather bothersome in elderly men and profoundly affect their quality of life (QoL) [1]. LUTS are generally defined in terms of 3 sets of symptoms: voiding, storage, and postmicturition [2]. Male LUTS often present as a result of benign prostatic hyperplasia (BPH), which involves increased resistance to urinary flow caused by bladder outlet obstruction, increased contractility of smooth muscle of the prostate and urethra [3], detrusor overactivity with denervation, decreased blood flow in the urinary bladder, and elevation of mediators (e.g., adenosine triphosphate and prostaglandins) [4,5]. Patients who are diagnosed with BPH are widely prescribed ╬▒1-adrenoceptor blockers, which can reduce voiding and storage symptoms (e.g., the former: slow stream, intermittent stream, and straining; the latter: daytime frequency, nocturia, and urgency). The mechanisms of action of ╬▒1-adrenoceptor blockers involve inhibition of the contractile response of smooth muscle in the prostate by endogenously released noradrenaline. In patients with BPH, ╬▒1-adrenoceptor blockers reduce prostate tone and bladder outlet obstruction to improve voiding symptoms [6,7], increase blood flow in the urinary bladder to improve storage symptoms [4], and inhibit the release of ATP from the urothelium [8]. Because ╬▒1-adrenoceptor blockers can lengthen micturition intervals, afferent sensory nerves are also considered a site of action of ╬▒1-adrenoceptor blockers [9,10].
Naftopidil ((┬▒)-1-[4-(2-methoxyphenyl)piperazinyl]-3-(1-naphthyloxy)propan-2-ol) is an ╬▒1D/1A-adrenoceptor blocker that improves LUTS suggestive of BPH. Naftopidil has been found to improve subjective complaints, objective symptoms, and QoL associated with male LUTS [11,12]. Naftopidil dosedependently inhibited phenylephrine-induced increases in prostatic pressure in canines when administered intravenously [13] and improved bladder overactivity and impaired bladder blood flow caused by bladder outlet obstruction in rats when administered orally [4]. In addition, naftopidil transiently abolished rhythmic bladder contraction in a dose-dependent manner in rats when administered intrathecally at lumbosacral levels [14]. In BPH patients, storage symptoms are much more bothersome to their QoL than voiding symptoms [1,15]. Suppression of the enhanced micturition reflex stabilizes micturition and the storage function of the urinary bladder.
Accordingly, we studied the mode of action of naftopidil on afferent nerve activity related to storage symptoms. In an experiment using rat spinal cord slices, 30 to 100 ╬╝M naftopidil facilitated miniature inhibitory postsynaptic currents (mIPSCs) in substantia gelatinosa (SG) neurons, as seen with the patchclamp procedure [16]. That study determined the mechanism of action of naftopidil on the micturition reflex and supported a previous study showing that naftopidil mediates gamma-aminobutyric acid (GABA)/glycine receptors at the level of the spinal cord [17]. However, the effect of naftopidil on excitatory postsynaptic currents (EPSCs) in SG neurons remains to be elucidated. The goal of this study was to further characterize the mechanism of action of naftopidil using the patch-clamp technique.
MATERIALS AND METHODS
All experiments were carried out in accordance with the Guiding Principles for Care and Use of Animals in the Field of Physiological Sciences of the Physiological Society of Japan and were approved by the local Animal Experiment Committee of Kumamoto Health Science University and Kyushu University. All possible efforts were made to minimize animal suffering and the number of animals used for the studies.
Preparation of Spinal Cord Splices
Spinal cord slices were prepared from adult male rats, and blind whole-cell patch-clamp recordings from SG neurons were performed as described previously in detail [18-20]. Briefly, Sprague-Dawley rats aged 6ŌĆō8 weeks were deeply anaesthetized with urethane (1.2 g/kg, intraperitoneally), and a lumbosacral laminectomy was subsequently performed. The lumbosacral segments of the spinal cord (L2ŌĆōS3) with ventral and dorsal roots were removed and placed in ice-cold Krebs solution equilibrated with 95% O2 and 5% CO2. The Krebs solution contained (in mM): NaCl 117, KCl 3.6, CaCl2 2.5, MgCl2 1.2, NaH2PO4 1.2, NaHCO3 25, and glucose 11 (pH, 7.4). After the spinal cord was isolated, the rats were immediately killed by exsanguination under urethane anesthesia. The pia-arachnoid membrane was removed after cutting all the ventral and dorsal roots, except for the L5, L6, or S1 dorsal root on 1 side, and any of them was used. The spinal cord was mounted on a vibratome, and a 550-╬╝m-thick transverse slice with the attached dorsal root was cut. The slice was moved onto a nylon mesh in the recording chamber in a volume of 0.5 mL of Krebs solution, and was completely submerged and perfused with Krebs solution saturated with 95% O2 and 5% CO2 at 37┬░C┬▒1┬░C at a flow rate of 10ŌĆō15 mL/min. The dorsal root was stimulated using a suction electrode (Fig. 1A).
Whole-Cell Patch-Clamp Recordings From SG Neurons
The SG was precisely identified with transmitted illumination as a relatively translucent band across the dorsal horn in the transverse slice preparations. Blind whole-cell voltage-clamp recordings were made of SG neurons, as described previously [16,20,21]. The patch pipettes were filled with a solution containing potassium gluconate solution (in mM): K-gluconate 135, KCl 5, CaCl2 0.5, MgCl2 2, EGTA 5, HEPES 5, and ATPMg 5 (pH, 7.2). The patch pipettes had a tip resistance of 6ŌĆō12 M╬®. Series resistance was assessed according to the response to a hyperpolarizing step of 5 mV. This value was monitored during the recording session, and data were rejected if values changed by >15%. Signals were acquired with a patch clamp amplifier (Axopatch 700A, Molecular Devices, Union City, CA, USA). The data were digitized with an AD/DA converter (Digidata 1321A, Molecular Devices), stored on a personal computer using a data acquisition program (Clampex version 9.0, Molecular Devices), and analyzed using a software package (Clampfit version 9.0, Molecular Devices). Cell recordings were made in voltage-clamp mode at holding potentials of -70 mV to record EPSCs [16,20]. To elicit EPSCs, the dorsal roots were stimulated at a frequency of 0.2 Hz (duration, 100 ╬╝sec). The A╬┤- or C-afferent-mediated responses evoked by dorsal root stimulation were distinguished based on the conduction velocity of the afferent fibers (C-fiber response: <0.8 m/sec; A╬┤-fiber response: 2.0ŌĆō11 m/sec) and stimulus threshold (C-fiber response: >200 ╬╝A; A╬┤-fiber response: 20ŌĆō200 ╬╝A), as previously described [19,20]. The conduction velocity was calculated from the latency of synaptic responses and the length of the dorsal root. The evoked responses were considered monosynaptic for the A╬┤ fiber-evoked EPSCs if the latency remained constant when the root was stimulated at 20 Hz, and for the C fiber-evoked EPSCs if no failure occurred regardless of the constancy of the latency when stimulated at 2 Hz [19,20]. To avoid the influence of evoked release on presynaptic terminals, dorsal root stimuli were not applied during the examination of the effects of drugs on spontaneous or miniature synaptic responses.
Drug Application
Naftopidil (PubChem CID: 4418) (Asahi Kasei Pharma Co., Tokyo, Japan) was dissolved in 1% dimethyl sulfoxide (DMSO) (PubChem CID: 679) (Wako, Osaka, Japan) in Krebs solution. The other drugs were dissolved in Krebs solution and applied by perfusion via a 3-way stopcock without changes in the perfusion rate or temperature. The other drug used in this study was prazosin hydrochloride (PubChem CID: 68546) (Wako).
RESULTS
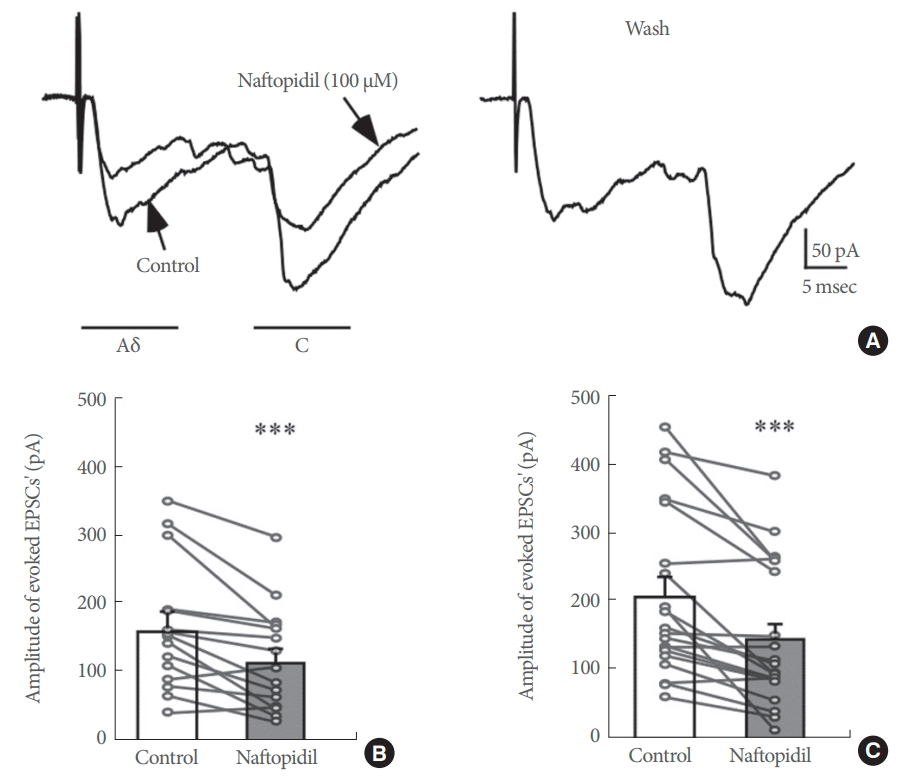
Monosynaptic EPSCs with short and/or long latencies were evoked in SG neurons by stimulation of the dorsal root attached to spinal cord slices. These 2 types of monosynaptic responses were considered to be A╬┤- or C-afferent-mediated responses based on the conduction velocity of the afferent fibers and stimulus threshold (Fig. 1B, C). The average conduction velocities of the afferent fibers were 6.5┬▒0.4 m/sec for A╬┤ fibers (n=15) and 0.58┬▒0.02 m/sec for C fibers (n=20). The threshold stimulus intensities for the monosynaptic A╬┤ fiber- and C fiber-evoked EPSCs were 0.10┬▒0.02 mA (n=15) and 0.91┬▒0.12 mA (n=20), respectively. These conduction velocities and stimulus intensities are comparable to those observed in previous studies [18,20,22].
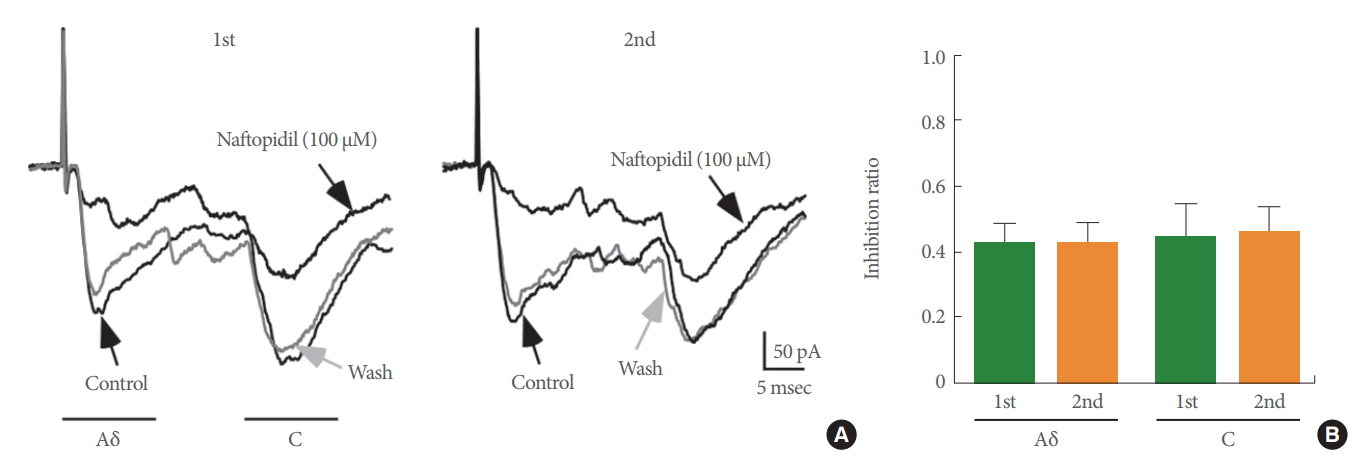
The effects of 100 ╬╝M naftopidil on monosynaptic A╬┤ fiberand/or C fiber-evoked EPSCs were investigated using 26 SG neurons in total (Fig. 2). Each of the evoked EPSCs was reversibly inhibited in amplitude by naftopidil. When examined in many neurons, the inhibition of the peak amplitudes of A╬┤ fiber- and C fiber-evoked EPSCs was 72.0%┬▒7.1% (P<0.001, n=15; Fig. 2A, B) and 70.0%┬▒5.5% (P<0.001, n=20; Fig. 2A, C), respectively. DMSO in which naftopidil is dissolved is toxic to cells [23]. To exclude the possibility that the observed effect was mediated by DMSO, we examined the effects of DMSO alone on these neurons. DMSO at 1% showed no effect when present in the perfusate of Krebs solution on any of the A╬┤ fiber- and C fiber-evoked EPSCs tested compared with the control (A╬┤ fiber-evoked EPSCs; 98.1%┬▒5.1% of control, P>0.05, n=2, C fiber-evoked EPSCs; 97.5%┬▒1.7% of control, P>0.05, n=4). These results are consistent with our previous studies [16]. Although enhancement of spontaneous inhibitory transmission, which is produced by naftopidil in SG neurons, is not subject to desensitization [16], we also examined how repeated application of 100 ╬╝M naftopidil affected A╬┤ fiber- and C fiber-evoked EPSCs. As shown in Fig. 3A, additional naftopidil was administered at an interval of 20 minutes. The first and second naftopidil treatments produced similar decreases in A╬┤ fiberand C fiber-evoked EPSCs (Fig. 3). Relative to the control, A╬┤ fiber- and C fiber-evoked EPSC amplitudes in the presence of the initial application of naftopidil were almost comparable to the amplitudes after the second application (Fig. 3B), indicating that naftopidil activity was consistent with recovery from desensitization. Because 100 ╬╝M naftopidil repeatedly suppressed A╬┤ fiber- and C fiber-evoked EPSC amplitudes, we next investigated how prazosin, another ╬▒1-adrenoreceptor antagonist, affected this suppression in the same neurons. Bath application of 10 ╬╝M prazosin had no effect on any of the A╬┤ fiber- and C fiber-evoked EPSCs compared with the control (A╬┤ fiber-evoked EPSCs: 97.2%┬▒1.8% of control, P>0.05, n=3; C fiber-evoked EPSCs: 97.2%┬▒5.1% of control, P>0.05, n=4) (Fig. 4).
DISCUSSION
To evaluate the effects of naftopidil, an ╬▒1-adrenoceptor blocker, on evoked EPSCs, patch-clamp recording was performed in SG neurons from the spinal cord of adult rats. Naftopidil inhibited the amplitude of both monosynaptic A╬┤ fiber- and C fiber-evoked EPSCs, and these effects were reversible and reproducible. Using the same procedure, prazosin inhibited neither A╬┤ fiber- nor C fiber-evoked EPSCs at all.
Mechanism of Action of Naftopidil
In this study, the reduction in evoked EPSCs in the spinal cord from L5 to S1 by naftopidil indicates suppression of nociceptive stimulation or the micturition reflex. In a previous study, naftopidil increased the frequency of mIPSCs, but not their amplitude [16]. The frequency of mIPSCs in the presence of naftopidil was attenuated with bicuculline or strychnine. These data indicate that naftopidil may act on the synaptic terminals of inhibitory interneurons and enhance the release of glycine and GABA. Therefore, naftopidil may activate inhibitory interneuron terminals in the spinal dorsal horn. The amplitudes of both GABAergic-evoked and glycinergic-evoked IPSCs were also enhanced by naftopidil [16]. This result suggests that naftopidil may change the sensitivity of GABA and/or glycine receptors in the spinal dorsal horn neurons. However, the amplitude of mIPSCs was not affected following bath application of naftopidil. Consequently, these results exclude the possibility that naftopidil changes the sensitivity of GABA and/or glycine receptors in the spinal dorsal horn neurons. In addition, naftopidil inhibits the micturition reflex, and intrathecal bicuculline and/or strychnine reverses the inhibition [17]. Therefore, naftopidil seems to elevate the levels of GABA and/or glycine at the presynaptic site of SG neurons in the spinal cord. Consequently, evoked EPSCs may be inhibited by a reduction in glutamate release together with upregulation of GABA and/or glycine levels by naftopidil in SG neurons. This could contribute, at least in part, to the inhibitory modulation of somatosensory sensation in the SG by intrathecally administered naftopidil.
Alternatively, naftopidil may inactivate the influx of cations into SG neurons, subsequently suppressing the release of glutamate. This possibility is supported by our previous studies, as follows. We examined the actions of naftopidil on miniature EPSCs in adult SG neurons and showed that naftopidil increased the frequency of miniature EPSCs [16]. In the present study, naftopidil blocked monosynaptic A╬┤ fiber- and C fiber-evoked EPSCs. Similar to the case of transient receptor potential channels in a previous report [20,22,24], we propose that naftopidil depolarizes the presynaptic terminals or axons of A╬┤ and/or C afferents by activating naftopidil binding sites (not ╬▒1D/1A-adrenoceptors) expressed on the presynaptic membranes. This depolarizing action has a facilitatory effect on miniature EPSCs and may also inhibit evoked release from A╬┤ and C fibers through inactivation of voltage-gated Na+ channels or by shunting of Na+ currents [25].
Regarding the negative effect of prazosin (10╬╝M), in addition to the findings of the previous studies mentioned above, naftopidil binds to human 5-HT2A and 5-HT2B receptors with pKi values of 6.55 and 7.82, respectively, and may functionally block the 5-HT2A and 5-HT2B receptors in the human urinary bladder [26]. However, outward current is induced in excitatory interneurons when 5-HT activates postsynaptic 5-HT1A receptors [27]. In contrast, in the small population of SG neurons, 5-HT induces an inward current mediated by the activation of postsynaptic 5-HT3 receptors on inhibitory interneurons [27,28]. Moreover, 5-HT inhibits glutamate release from C afferent fibers by activating presynaptic 5-HT1A-like receptors [29]. However, 5-HT1A, 2A, 2B, 2C, 3 receptor antagonists do not affect the excitatory and inhibitory synaptic response [27,29,30]. These results suggest that naftopidil must have acted on a binding site that does not bind prazosin. This binding site remains to be identified. However, previous research found that GABA and/or glycine receptors mediated the facilitation of mIPSCs and evoked IPSCs by naftopidil [16]. Therefore, GABAergic or glycinergic neurons may inhibit excitatory glutamatergic neurons. Finally, naftopidil may decrease excitation of the micturition reflex through such neural networks (Fig. 5).
Reversibility and Reproducibility
The effects of naftopidil on evoked EPSCs were reversible, suggesting that naftopidil may act on a binding site, likely its receptor. However, because prazosin did not inhibit evoked EPSCs, the effect of naftopidil on evoked EPSCs may not involve ╬▒1-adrenoceptors. In addition, naftopidil reduced evoked EPSCs in a reproducible manner, and therefore, naftopidil is not considered to produce desensitization of SG neurons, even at a high concentration.
Concentration of Naftopidil
Concentrations of naftopidil from 0.3 to 3 ╬╝M, 0.4 to 40 ╬╝M, and 0.5 to 100 ╬╝M have been used to determine detrusor contractility [26], platelet aggregation responses [31], and the viability of cancer cells [32,33], respectively. In the present study, a high concentration of 100 ╬╝M was used, although this concentration has also been used in previous studies. This concentration of 100 ╬╝M seems to be above that used clinically. However, the present results are probably useful for future studies, such as pharmacological studies of structure-activity correlations, the development of new drug delivery systems for local injection, and the development of derivatives with higher activity.
In conclusions, the present study suggests that naftopidil reduces the amplitude of evoked EPSCs, although its mechanism of action apparently does not involve ╬▒1-adrenoceptors. In addition to the facilitation of mIPSCs, as reported in a previous study, the inhibition of evoked EPSCs by naftopidil may also contribute to suppression of the micturition reflex together with nociceptive stimulation. Because these responses seem to be associated with analgesia, naftopidil or more highly active derivatives may be useful for treating LUTS with concomitant pain, such as interstitial cystitis/bladder pain syndrome [34].